数据 / 观察资讯
全球儿童微生态研究进展(2018-2019)
2019-09-29 22:47 来源:新营养


深圳市微健康基因研究院授权发布
免责声明
儿童微生态研究作为当前儿科医学领域最为重要的热点之一,已受到越来越多的临床医生和科研专家的关注。本报告就近年来部分儿科疾病微生态的突破性研究进行了整理总结,文中的信息都来源于公开发表的文章、政府公告等。
因目前儿科微生态研究所涉及领域较广,材料收集和编撰时间较为仓促,部分研究成果及进展未在文中列出,恳请各位专家批评指正,也欢迎各位专家提供专业意见和建议:zhouqian@wehealthgene.com。
版权声明
本报告版权属于研究团队共有,研究团队保留追究法律责任的权利,其他人员未经许可不得擅自使用。
研究团队(以姓氏拼音为序)
戴文魁 深圳市微健康基因研究院
葛 兰 华大精准营养
黄志华 华中科技大学同济医学院附属同济医院
栗东芳 深圳市微健康基因研究院
刘 盈 首都医科大学附属北京儿童医院
马 琳 首都医科大学附属北京儿童医院
舒赛男 华中科技大学同济医学院附属同济医院
王文建 深圳市儿童医院
武庆斌 苏州市儿童医院
杨振宇 深圳市微健康基因研究院
张 琳 河北医科大学第三医院
郑跃杰 深圳市儿童医院
周 潜 深圳市微健康基因研究院
校对、排版
周潜、黄惠萍 深圳市微健康基因研究院
一、儿童微生态与疾病研究进展
(一)肠道共生菌群与食物过敏症
人体消化道黏膜聚集了数以几十万亿计的微生物群落,它们利用宿主分泌物、肠上皮脱落细胞、食物残渣及其他微生物提供的营养素,通过菌体直接作用,或者释放代谢产物间接作用于宿主粘膜和/或正向/ 反向影响其他微生物,共同组成了互惠互利、互相依存的共生体系。
人体免疫系统能够耐受这些微生物而不产生排斥性免疫应答,也避免对致病菌多度应答造成宿主局部组织损伤。
介导宿主-菌群共生体的机制高度复杂,需要频繁互作协调。肠道菌群结构和/或代谢产物发生改变,可打破宿主与共生菌群之间的协调关系,造成宿主免疫稳态机制破坏,宿主对环境因素产生过度免疫应答(超敏感)。
食物过敏症是由饮食、肠道共生菌群及其宿主相互作用形成的粘膜免疫耐受发生偏移而对食物抗原产生不适度的免疫应答性疾病。
越来越多研究显示肠道菌群是导致食物过敏症发生的关键因素,肠道微生态、饮食、宿主基因与食物过敏症存在强烈的内在联系。
1、研究比较食物过敏症患儿差异化菌群
牛奶过敏症治愈婴儿肠道有特定的菌群结构, 表现为肠道富集梭菌科和厚壁菌门。
隶属于毛螺杆菌科和瘤胃球菌科的某些菌属与鸡蛋过敏症相关,生命早期(婴儿期)肠道菌群与 8 岁时鸡蛋过敏症治愈无关;
一项宏基因组关联研究发现,Haemophilus, Dialister, Dorea 和Clostridium 菌属在食物高敏感儿童肠道定植丰度低,而 Citrobacter, Oscillospira, Lactococcus 和 Dorea 菌属在食物过敏症儿童肠道定植丰度降低;
Feehley 等研究证实牛奶过敏症婴儿粪便可导致无菌鼠发生食物过敏症,而健康婴儿粪便具有保护无菌鼠抵御食物过敏症的作用;
他们结合宏基因组学、转录组学和代谢组学技术,不仅找出具有抗食物过敏症的婴儿肠道菌群——粪厌氧棒状菌(Anaerostipes caccae,它是属于厚壁菌门下、梭菌目、毛螺杆菌科下的一个多糖发酵菌种),还将分离提取的粪厌氧棒状菌在无菌鼠上模拟出抗过敏症转录组和表型。
这项发现为临床食物过敏症早期诊断、干预策略和效果评价,提供了又一个重要的肠道菌种标识物。
2、肠道菌群可能是治疗食物过敏症的重要靶点
生命早期(新生儿期、婴儿期)膳食和肠道菌群驱动着婴儿免疫系统成熟发育,尤其是肠道菌群结构和代谢物活性通过调节宿主肠粘膜免疫,诱导了口服耐受的建立,分子机制在于:肠道菌群通过诱导RORγt Treg 细胞和 Th17 细胞发挥调控 Th2 应答的作用。
激活树突细胞表面的模式识别受体是肠道菌群独有的促进 Treg 分化成熟的路径。因此,肠道菌群失调,尤其是生命早期肠道菌群失调可显著提高过敏性疾病的易感性。
母乳寡聚糖(HMOs)选择性促进肠道保护性菌群双歧杆菌增殖,帮助婴儿肠道菌群建立和成熟。不同母乳寡聚糖结构与食物过敏症存在关联,可能机制是:不同 HMO 结构谱分别靶向支持婴儿肠道不同菌种增殖,间接影响着婴儿期免疫系统发育成熟和后天教育。
总之,目前大量人群研究提示:食物过敏症可能与生命早期肠道菌群暴露有关,后者修饰了宿主免疫系统发育,或促进了宿主对食物抗原产生病理性免疫应答。
3、口服免疫疗法(OIT)成为临床治疗食物过敏症的有效手段
肠道菌群通过诱导早期宿主口服耐受建立和免疫系统发育成熟,在后续食物过敏症发生发展中发挥关键作用,因此,生命早期(婴儿期)是干预的“关键窗口期”。
饮食是调节肠道菌群结构和功能的最关键的手段,英国三项高质量临床研究证明,婴儿期主动摄入花生显著降低儿童期过敏症发生率,2017 年美国多家专业学会颁布预防花生过敏症补充指南:推荐生命早期适当摄入含花生食物初级预防儿童期花生过敏症。
但是另有研究发现,4-6月龄中重度湿疹婴儿或健康婴儿接受 4-6个月的全鸡蛋干预,对1岁时鸡蛋过敏症发生率的干预结果并不一致。
未来还需要对食物过敏症发病机制和影响因素做更深入的研究,以及临床干预试验的不断丰富。
4、口服益生菌预防/治疗食物过敏症需要进一步验证
LGG联合深度水解蛋白奶粉可提高6-12月龄婴儿牛奶过敏症的持续无应答率;
花生过敏症儿童连续18个月接受LGG联合花生口服免疫疗法(OIT),联合治疗组儿童持续无应答率显著高于单用 OIT 治疗组,此外,免疫学指标也发生显著改善(如降低特异性 IgE、提高特异性 IgG1),试验结果提示益生菌对 OIT 治疗花生过敏症儿童具有增效作用;
该研究团队继续干预至 4 年,加用益生菌膳食干预组患儿在持续无应答、皮刺风疹直径以及特异性抗原水平等临床表型和实验室表型方面,过敏症状获得持久改善;
这项临床研究还需要进一步明确单用LGG对比OIT疗法对食物过敏症的疗效权重,因为该研究缺乏单用LGG对照组。
数项研究结果提示,婴儿期益生菌干预降低食物过敏症风险的未来前景看好,现有研究资料尚不足以支持膳食益生菌可改善食物过敏症的结论,还需要进一步积累证据。
粪菌移植(FMT)用于食物过敏症的临床研究尚未见到文献报道, 在美国临床试验注册中心(https://clinicaltrials.gov)网站上查询,粪菌微胶囊治疗成人花生过敏症的临床1期试验、以及益生菌预防婴儿湿疹和食物过敏症正在进行临床受试者招募。
5、目前的挑战和机会
a) 目前所有研究都是排他性地聚焦在细菌微生态,忽略了可能同样重要的真菌生态组和病毒生态组,描述真菌组和病毒组的研究方法学还在开发中;
b) 鉴于食物过敏症病因和发病过程的复杂性,多模型多手段共同预防和/或治疗食物过敏症,要比单一的手段更有效;
c) 对肠道微生态在皮肤湿疹和呼吸道哮喘发病中作用缺乏深入研究和理解;
d) 以肠道菌群为靶点的饮食免疫疗法已经有明确的实质性进展,在湿疹或哮喘治疗上还没有出现被证明确实有效的免疫疗法或生物活性治疗手段。
参考文献21篇(略)
(二)儿童肠-脑轴发育及神经发育性疾病(自闭症谱系障碍)
“脑-肠”介导了肠道及共生菌群与神经系统的双向沟通,涉及肠道菌群代谢物、宿主神经、内分泌、代谢和炎症机制;
肠道菌群在调节情感、社交、疼痛感知和消化行为方面发挥重要的作用,这种互作的改变可能参与了几种中枢-肠道疾病的发生发展;
生命早期神经系统发育伴随着脑-肠互作的程序性建立, 这个时期暴露各种环境因素不仅塑造肠道菌群器官发育,影响着肠道菌群与神经系统互作关系,也为各种胃肠道和神经系统发育性疾病的发生埋下伏笔;
大量研究证据支持肠道菌群可能在自闭症( ASDs)发生上发挥着协同促进作用,ASD 患儿存在广泛的肠道菌群紊乱,而肠道菌群紊乱法反过来促发了 ASD 患儿的全身低水平炎症和代谢紊乱;
益生菌在治疗神经行为障碍和肠道功能紊乱方面非常具有前景,现有临床依据还很有限,且异质性高,未来需要设计良好的随机对照临床试验验证益生菌对 ASD 的有效性。
1、“脑-肠轴”互作的信号通路和分子机制
“脑-肠轴”定义为肠道菌群通过神经、内分泌和免疫路径与中枢神经系统之间建立起来的沟通交流的生理性网络。这种沟通是双向的,大脑对肠道菌群的作用以及肠道菌群影响大脑和行为是同等重要的。
肠道菌群代谢物承担了“脑-肠轴”自下而上的信号通路。
最好的例证就是短链脂肪酸( SCFA)、次级胆汁酸(2BAs)和色氨酸代谢物,这些代谢产物直接作用于肠道内分泌细胞、肠道嗜镉细胞和粘膜免疫系统,自下而上地传导信号,另外,其它肠菌代谢物能够通过肠屏障进入全身循环,甚至通过血脑屏障进入脑组织。
目前还不清楚肠道菌群代谢物到达中枢神经系统并修饰远处大脑的详细通路和分子机制, 也或者微生物信号通过迷走神经和/或脊髓传入神经通路与中枢进行互动交流。表1汇总了肠道菌群代谢物影响神经系统的信号通路及效应。
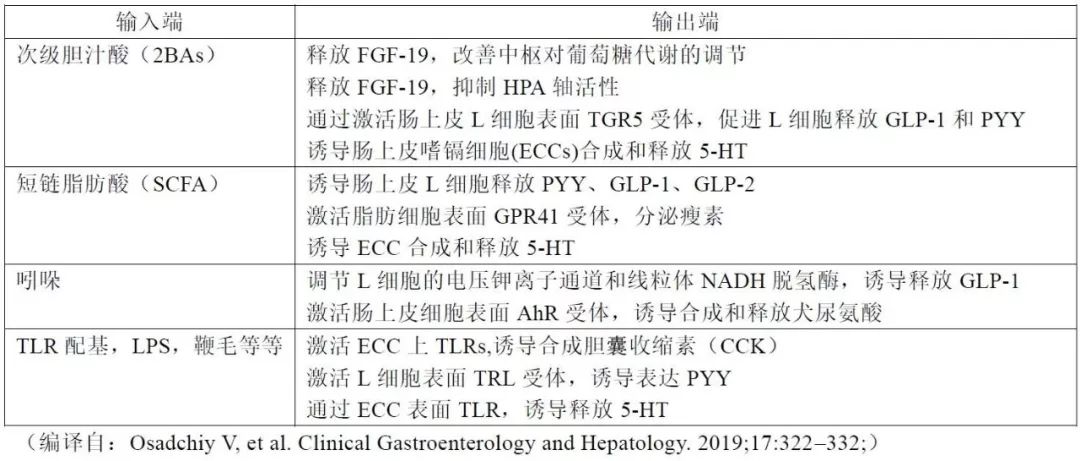
表1 肠道菌群代谢物到中枢神经系统的信号通路
“脑-肠轴”自上而下的信号通路是指中枢神经系统(CNS)向肠腔释放神经内分泌介质,大脑通过各种方式支配或影响肠道菌群结构和功能。
压力暴露是CNS影响肠道菌群的主要机制,母子分离实验表明,母亲产后压力与肠道乳杆菌丰度显著减少相关,这导致肠道菌群结构紊乱,并伴有明显压力行为,子鼠表现为对机会性感染的易感性增高。
CNS的直接信号包括儿茶酚胺(受躯体和精神压力影响),间接信号包括自主神经系统(ANS)的两个分支(交感神经系统和副交感神经系统),ANS诱导肠道菌群发生生理变化,后者影响肠道菌群结构和功能。
例如,改变肠蠕动频次会影响水分含量、营养素供应和细菌清除率,肠蠕动动力减弱导致细菌过度增殖,肠道传输时间延长与粪便菌群丰度和结构高度相关。
ANS通过调节肠壁杯细胞分泌和肠上皮细胞内通透性影响肠粘膜完整性,在小鼠脑损伤模型中发现去甲肾上腺素释放增高,杯细胞数量降低,粘蛋白分泌减少,综合作用导致肠道菌群结构变化,这种变化与脑损伤程度呈正相关;
大多数脑损伤和“脑-肠轴”疾病都表现为压力应答增高以及ANS功能改变,CNS对肠道菌群的自上而下的调节作用在肠道菌群变化中发挥重要作用。
2、生命早期肠道菌群影响大脑程序性发育
生命早期(0-3岁)不仅是中枢神经系统(CNS)建立广泛的神经元突触链接和髓鞘生成的重要时期, 也是肠道菌群结构建立和功能成熟的时期;有研究者认为产前胚胎期也是肠道共生菌群的一个重要的发育时期,微生物获取热量对于大脑发育至关重要。
动物研究提示,生命早期肠道菌群对于调控前额叶皮层的髓鞘形成发挥重要作用,无菌鼠编码髓鞘结构成分的基因上调,导致出现超髓鞘化。
对89例婴儿的随访研究提示,1岁时婴儿肠道共生菌群结构与1岁后的认知能力相关,证实了生命早期肠道菌群与大脑发育之间的互作关系。
最近研究发现,肠道菌群代谢产物还参与了宿主CNS的发育和功能维持,生命早期缺乏肠道菌群定植,提高了大脑杏仁核的转录表达,后者参与大脑情感、焦虑和社交行为。
生命早期各种环境微生物、饮食、抗生素、压力和其它因素塑造了肠道菌群结构和功能,影响着肠道菌群与快速发育的CNS之间的互作,这也意味着不良环境因素暴露为各种神经发育性疾病的发生发展埋下病因。
阴道产婴儿肠道定植菌群与母亲阴道和粪便菌群相似(富含乳酸杆菌和普雷沃杆菌属);母亲阴道菌群是动态变化的,受产妇精神压力的影响,研究证实:母亲阴道菌群不仅影响新生儿肠道菌群结构,也影响着肠道代谢组;孕妇压力(感染、心理社会压力)可增加新生儿对精神分裂症、自闭症、注意力缺失的易感性。
剖宫产新生儿肠道富集葡萄球菌属和棒状杆菌属,尤其是阴道产婴儿和剖宫产婴儿在皮肤、肠道和鼻咽部菌群结构上的差异一直持续到出生后2岁,而这个时期又是CNS深入发育的最关键期。
一项200万丹麦足月儿的出生队列研究发现,剖宫产与儿童期哮喘、炎症性肠病、免疫缺陷和其他慢性免疫性疾病发生率升高相关。
肠道菌群还受到孕期母亲营养状态以及产后出生后喂养方式的影响。母乳喂养婴儿的神经发育优于奶粉喂养婴儿,其肠道富含高丰度多样化的双歧杆菌属,母乳中含有构建婴儿肠道菌群最关键的因素,即母乳寡聚糖(HMOs),这些大分子糖类无法被婴儿上消化道吸收利用,而被结肠菌群发酵利用,促进了肠道菌群发育。
人群研究发现,母亲在孕期摄入高脂饮食改变了新生儿共生菌群,可能与儿童期自闭症(ASD)高易感性相关,持续母乳喂养6个月与ASD低易感性相关。
小鼠模型研究发现,给孕鼠鼻饲抗癫痫药丙戊酸(VPA)可诱发出子鼠自闭症样社交障碍和肠道厚壁菌门和拟杆菌门丰度变化。
3、肠道菌群变化与ASD相关胃肠道和免疫功能障碍之间存在关联
自闭症谱系障碍(ASD)是一种病因不明的、病理生理学模糊的、严重的神经发育性疾病,临床表现为认知能力低下、沟通障碍、重复行为、社交和沟通障碍导致患者在社会上处于弱势局面。
与其它神经发育性疾病相比,自闭症患者的合并症最为神秘莫测,其合并症造成的负担最高,常见的合并症包括癫痫发作、精神疾病和胃肠道症状。
研究提示ASD患者肠道菌群结构和功能发生了显著变化,ASD患者粪便菌群富集梭菌属,拟杆菌/厚壁菌比值降低,乳酸杆菌属和脱硫弧菌属丰度升高,并与ASD严重程度相关;ASD患者粪便SCFA(乙酸、丙酸、丁酸)水平显著降低,与ASD病情相关。
ASD患者中一些对碳水化合物代谢至关重要的菌属(如普雷沃杆菌属、粪球菌属、韦荣球菌属)丰度显著降低;
另一方面,取ASD患儿十二指肠粘膜进行活检发现,上消化道萨特氏菌属丰度显著升高,该菌属参与粘膜降解代谢,患儿治疗后胃肠道和神经行为症状再次复发,推测原因肠道梭菌目对万古霉素耐受,进而过度繁殖转成感染型。
另外,梭菌科也产生释放一些代谢产物(如酚类、对甲苯酚和吲哚衍生物),对人体具有潜在毒性,在自闭症患儿的粪便中可检出上述代谢产物。
虽然ASD患者肠道菌群发生改变,但是目前还无法定义ASD的肠道菌群特征,原因是受试者的高度同质性,如年龄、饮食、药物治疗、区域、合并症以及神经行为和胃肠道症状严重程度。
未来研究要充分考虑各研究受试者的异质性,正确评估ASD患者肠道菌群变化的研究结果。最近一项研究推测甘草膦(GLY,一种环境杀虫剂)通过促进产毒素梭菌目增殖加重自闭症病理过程。
产气荚膜梭菌和肉毒梭菌都对GLY 高度耐受,而其它肠道有益菌如双歧杆菌和乳酸杆菌对GLY敏感,因此环境GLY可能会恶化肠-脑轴互作,通过改变肠道菌群结构释放梭菌目毒素,加重自闭症病理。
肠道菌群与免疫系统表现出重要的双向相互作用,ASD患者的许多免疫指标发生失调,循环系统和神经系统中的细胞因子、化学因子和其它炎症因子,以及各种细胞亚群的异常分布和高反应,这些ASD相关的免疫异常是最近研究的热点。
许多免疫表型与升高的促炎症状态高度一致,主要是指升高的细胞因子和化学因子,包括IFN-γ、IL-1β、IL-6、IL-12p40、TNF-α、单核细胞趋化蛋白-1、转化生长因子β、趋化因子配体2,以及过度活跃的细胞免疫应答。
ASD患者表现出不同的免疫异常,包括免疫细胞/细胞因子谱的差异性变化,还难以确定ASD个体免疫与肠道菌群之间的直接联系,另外饮食、生活方式和遗传等混杂因素也会修饰免疫应答。
总之,目前还不能确定共生菌群紊乱与ASD个体免疫失调之间的因果关系,还需要更多的研究以确定ASD中共生菌群变化是否能充分引起任何一种ASD的免疫异常表型。
自闭症儿童消化道症状与炎症状态有关,几项最新综述也指出合并消化道症状的ASD患儿存在着炎症和免疫失调。Boso等提出自闭症儿童存在的“炎症假设”,即AGE-RACE轴功能失调,表现为全身炎症,高水平RACE和促炎性配基S100A9升高。
最新一项研究发现, ASD患儿的星形胶质细胞表达更多的促炎症因子,可能促进了ASD患儿神经元发育和突触形成发生偏移。
另外一种观点认为肠屏障功能障碍导致肠道通透性增加,而肠道菌群及代谢产物对于维持肠屏障功能至关重要,而ASD患儿肠屏障功能障碍增加了肠粘膜通透性。
这种状态也称作“肠漏”,导致大量肠道细菌、毒素如LPS和代谢产物进入体内激活免疫系统,诱导炎症因子进入体循环,激活的免疫系统释放大量炎症因子和化学因子,后者调节CNS影响大脑发育早期阶段,促进了自闭症病理过程。
4、肠道菌群是ASD危险因素的潜在媒介
肠道菌群处于基因与环境的交汇界面,肠道菌群的结构和功能不仅取决于遗传背景,并受到年龄、感染、饮食和异生质等环境因素的重要影响。
生命早期共生菌群改变不仅对该个体健康和疾病产生持久影响,还将感染的持久影响传递给该个体,调节宿主基因组的表观遗传修饰。
有意思的是,共生菌群诱导的表观修饰可以决定宿主的转录谱,例如,短链脂肪酸的丁酸可抑制组蛋白去乙酰化,后者参与了细胞周期进展、基因沉默、分化和遗传毒性反应。
目前还不清楚是否肠道菌群介导了遗传或环境风险因素促进ASD发生发展。但是越来越多的证据表明,肠道菌群变化应对了ASD的病因危险因素。
基于大量流行病学、人群和动物研究发现,产妇感染是泛自闭症的主要环境危险因素,模拟母体免疫激活的子代鼠成年后肠道菌群结构发生普遍改变,共生菌群紊乱与顽固性行为异常、神经病理、免疫功能障碍和缺乏胃肠道屏障完整性高度相关。
孕期暴露环境危险因素或遗传风险而导致孕母共生菌群改变可以传递给后代,母亲或产前暴露遗传或环境危险因素是疾病发生的原因。大量证据表明哺乳动物胎儿通过分娩过程继承了上一代的共生菌群。
不论是自然阴道分娩或是剖宫产分娩都推动了子代菌群最初的播种和定植,阴道产婴儿的共生菌群与剖宫产婴儿的差别巨大,这一点对ASD等发育性疾病意义重大。
5、肠道菌群对ASD影响的未来研究方向
a) 新的研究显示,肠道菌群是胃肠道生理、免疫功能和大脑行为的重要调节剂,ASD患儿合并了胃肠道生理、免疫功能以及大脑行为的异常症状,需要更深入地分析和描述这些合并症状,以明确这些合并症与ASD关联的性质、强度和可重复性。
b) 对遗传背景、病史和ASD严重程度以及其他因素的评估有助于理解某种特定ASD亚型与特定症状的关联,帮助建立肠道菌群紊乱、胃肠道功能失调或免疫失调可能促进或恶化ASD行为异常的假设推断。
c) 利用新的和现有的动物模型,进一步研究ASD合并肠道菌群特征、胃肠道和免疫症状,重点分析不同遗传背景和环境ASD风险模型之间的共聚表型特征。
d) 需要进一步研究证实共生菌群紊乱、消化道功能或免疫应答异常能充分导致ASD的初级行为特征。从悉生动物或异生动物的研究开始,确定外周靶点和大脑的特定变化,开发新的ASD治疗策略。
e) 实施粪菌移植(FMT)对ASD疗效的高质量临床研究,为ASD人群提供所需的治疗指导。
参考文献23篇(略)
(三)肠道菌群对儿童生长发育的作用
生命早期共生菌群的建立和成熟对于个体免疫、内分泌、代谢和其它发育方面发挥着关键性作用。
这个时期各种环境因素暴露(饥荒和感染)可扰乱正常微生态的程序性建立,导致个体终身成长发育受影响和几代人的发育缺陷。
有关人类生命最初1000天(指的是从受孕到2岁的时期)的共生菌群建立和演替对个体生长发育作用的研究进展,提示我们:“营养不良”共生菌群可通过代际进行传播,导致个体成长发育损伤延续到下一代;
生命早期是对紊乱共生菌群进行恢复和矫正的最佳时间窗,以肠道共生菌群为靶点的干预手段有望成为预防和治疗儿童生长发育缺陷的临床新策略。
关于人体共生菌群初始化定植是胚胎期还是出生后的话题还在争论中。
最近几项出生队列研究间接提示,宫颈处的某些细菌有可能随着受精过程与精子一同进入子宫,并在受精、植入和早期胚胎发育时进入卵子内部,研究也证实了体循环血液或胎盘组织中存在细菌 DNA;
但是主流观点仍然认为:由于人体免疫系统的存在,子宫、胎盘、胎儿以及血液不可能存在共生菌群,胎儿发育期是无菌的概念会沿用下去。
最新一项研究采用四种方法(培养法、实时定量PCR、16S rRNA和宏基因组)检测足月产孕妇的胎盘样本,证实足月产孕妇的子宫是无菌的。
定义“正常”或“健康”的肠道共生菌群面临挑战,肠道菌群成熟度概念及共生菌群-年龄的z值(WAZ)的提出,是对儿童期生长发育评价体系的有力补充。
婴儿期0-6个月内最具年龄甄别效力的菌种包括长双歧杆菌和嗜热链球菌,而柔嫩梭菌、瘤胃球菌和Dorea菌(Dorea longicatna and Dorea formicigenerans)是6-24个月期间健康婴幼儿最具年龄甄别效力的肠道菌种。
1、人体微生态菌种/株通过人类世代传播,母婴垂直转移是最原始的机制
母体微生态通过母亲因素(如免疫应答力)或母体共生菌群代谢物通过胎盘间接影响胎儿发育,其它因素如饮食、压力或神经内分泌不仅直接影响母体微生态,还间接影响胎儿的表观遗传。
怀孕期间母亲肠道、阴道微生态随着孕期发生变化,这些变化是否对母亲或婴儿有适应性价值仍是未知数。
胎儿分娩代表着子代第一次暴露于外部环境并开始接触复杂的微生物,也是哺乳类动物一代又一代传递微生物菌种的原始机制。
胎生哺乳动物进化出了用于生殖的独立腔道(阴道)、供排泄的肛门和尿道,产道总是与直肠相邻但不与尿道相邻, 这提供了阴道和肠道发生微生物的代际传递的有效机制,羊膜绒毛膜的破裂让胎儿暴露于母亲的阴道菌群和四周粪便菌群。
新生儿出生时全身皮肤和口腔天然地暴露于母亲阴道共生菌群,并将这些微生物吞咽下去。因此,我们每个个体都从其母亲和外祖母身上继承了最初始的微生物群,就这样通过母系进行垂直传递,菌群可追溯到更早的人类祖先。
母乳是由复杂生物配方组成的单一食物,不仅富含新生儿生长发育所需的全部营养素,还有独立的微生态,为新生儿肠道共生菌群的建立提供了巨大的菌种来源;
母乳寡聚糖对婴儿肠道菌群的构建施加选择性压力,母乳中不同类别的碳水化合物可塑造婴儿肠道菌群和母乳微生态菌群,尤其是双歧杆菌菌种的丰度,新生儿在绝对母乳喂养期间,不仅大脑感知力和活力得到大幅度快速发展,还降低晚期罹患免疫性疾病的风险。
近期几项出生队列研究认为,生命早期共生菌群菌种来源于母亲相关菌种和环境菌群,其中母体微生态(肠道、口腔、阴道、皮肤)是子代共生菌群建立的最大贡献者,母体来源的菌株选择性而持久传递给阴道产婴儿的肠道,极少被环境的菌株替代。
2、母体共生菌群影响孕期胎儿的发育
母体阴道菌群对足月阴道产婴儿的肠道菌群定植影响很大。
母亲阴道微生态与子宫内胎儿形成互作关系,影响产前胎儿的程序性发育。当孕妇发生阴道炎时,致病菌上行性入侵子宫,刺激子宫形成炎症瀑布,导致小于胎龄儿(SGA)和早产儿。
阴道菌群的模式特征也与胎儿发育有关;在一项高收入国家城市出生队列研究中,孕妇的阴道菌群通常以四种乳酸菌中的一种为优势菌,绝大多数马拉维农村女性阴道菌群属于缺乏乳杆菌的多菌种优势。
将乳杆菌缺乏、多种菌优势的阴道菌群细分成四种类型,其中有一种类型由普雷沃杆菌属、孪生球菌属和棒状杆菌属占优势,这种类型阴道菌群与新生儿低LAZ值(身长-年龄Z值)高度相关,低LAT新生儿部分源自早产。
怀孕期间母体其它部位的共生菌群也影响着胎儿发育和妊娠结局。
母亲罹患炎症性肠病(IBD)与早产和低出生体重(LBW)相关;在高收入国家,早产还与母亲肠道菌群多样性降低以及低丰度的双歧杆菌属、链球菌素和梭菌属相关。
几项研究观察到母亲口腔微生态与胎盘和婴儿口腔微生态高度相似,提示母亲-胎儿之间存在共生菌群传递。
孕期母亲牙周感染合并口腔菌群紊乱与早产和SGA高度相关,提示母亲口腔共生菌群影响发育中的胎儿。
新生儿出生体重与母亲唾液中奈氏放线菌丰度呈负相关,而与干酪乳杆菌丰度呈正相关;在中低收入国家,由于口腔卫生差,孕妇患牙周疾病发生率高,她们早产和LBW发生率是高收入国家的4-5倍,母亲孕期牙周感染引起子宫内慢性炎症环境,而导致早产和SGA,或者婴儿发育不良。
3、母乳喂养深刻影响婴儿成长发育和肠道共生菌群程序化建立
健康足月新生儿肠道共生菌群高度接近母亲粪便、阴道或皮肤共生菌群,出生方式是主要影响因素,首先定植于婴儿肠道的菌种是典型的兼性厌氧菌,随后在6个月内逐步植入专性厌氧菌包括双歧杆菌、拟杆菌属和梭菌属。
据估计婴儿肠道有20%-30%菌种来自母乳,生命早期肠道菌群对内分泌、粘膜免疫和中枢神经系统发育成熟发挥重要作用,可能的机制是:共生菌群驱动胰岛素样生长因子1(IGF-1)在早期生长发育中发挥关键作用。
母乳在婴儿共生菌群建立中发挥重要作用,早产儿肠道菌群建立和演替可通过母乳喂养获得修饰,那些母乳喂养的早产儿出院时生长发育指标要显著优于奶粉喂养早产儿。
母乳喂养通过修饰婴儿肠道菌群直接促进了生长发育,母乳内存在独立的共生菌群,母乳菌群结构和功能随着母亲体重、出生方式、种族而变化,最常见的是变形菌门(主要是假单胞菌属)、链球菌属和葡萄球菌属,菌种主要来自母体皮肤、口腔和肠道共生菌群。
母乳菌群与婴儿生长发育之间的关系还未见研究报道。
母乳寡聚糖(HMO)在定义早期生长发育表型中起着重要作用。遗传因素影响母乳HMO成分,即活性岩藻基转移酶2(FUT2)的基因载体,可促进产生更多的岩藻糖基化和唾液酸化HMO,母乳的分泌状态影响了婴儿肠道共生菌群结构,分泌型母乳喂养婴儿的肠道更富集双歧杆菌属。
不论是高收入国家还是低收入国家,母乳HMOs都与婴儿生长和体格发育高度相关。
Charbonneau等分析发现马拉维发育迟缓婴儿的母亲表现为6个月时母乳唾液酸化HMO含量显著降低,将发育迟缓婴儿的粪便菌群移植给无菌鼠,之后给予“马拉维饮食”。
当喂饲了与母乳HMO结构类似的牛乳寡聚糖后,该组小鼠体重和骨体积明显增加,而对照组小鼠则没有表现出生长促进效应,提示母乳HMO促进生长作用具有菌群依赖性。HMO通过塑造婴儿肠道共生菌群介导了个体生长发育。
4、肠道共生菌群持续地影响婴幼儿的生长发育
母乳喂养结束后,引入固体食物促发了婴儿肠道菌群结构和功能多样性的大幅快速发育,肠道共生菌群成熟度上接近成人水平,成熟的肠道共生菌群优势菌属主要是降解多糖、粘液和复杂碳水化合物、并释放短链脂肪酸(SCFA)。
6个月到2岁是儿童生长发育呈线性发育的关键时期。
营养不足(定义为LAZ< -2)是全球最常见的营养不良表型;营养不良婴儿肠道缺乏长双歧杆菌和粘液乳酸杆菌,而富集脱硫弧菌属,肠道共生菌群多样性降低、氨基酸球菌属丰度升高,肠道富集谷氨酸发酵路径。
研究发现营养不良婴儿粪便中来自口咽部的菌种增多,丰度显著升高,提示营养不良婴儿消化道的区隔消失了,口咽菌群定植在下消化道部位,与生长发育障碍及全身炎症相关。
急性重症营养不良(SAM)和营养不足都与宿主代谢紊乱的表型相关,尤其是能量代谢、营养素代谢、氨基酸转运;小鼠断乳后喂饲缺乏锌和蛋白的饮食引起小肠菌群结构显著改变,并伴随能量代谢紊乱和膳食胆碱路径上调,经过营养膳食治疗后也无法恢复,表明营养不良可造成肠道菌群代谢持久性紊乱。
营养不良的巴西儿童尿液富集苯乙酰谷氨酰胺(PAG)、4-甲苯基硫酸盐(4-CS)、3-吲哚硫酸盐(3-IS),这些分别是氨基酸苯基丙氨酸、酪氨酸和色氨酸的肠道菌群代谢物,必需氨基酸减少与宿主肠道菌群紊乱相关,营养不良极有可能是通过宿主肠道降解蛋白的菌群介导发生的。
最近一项研究提示,宿主代谢紊乱是由共生菌群功能和结构变异导致的,主要是获得了部分菌种的功能基因,参与脂肪酸和氨基酸代谢的宏基因组路径在营养不良的个体内丰度显著不足,宏基因组参与修饰了个体的营养状况。
共生菌群对个体生长表型的影响是通过间接影响内分泌轴而发挥作用的。IGF-1是生长激素(GH)的介质,无菌小鼠中IGF-1显著耗竭,这种效应在断奶后更加恶化,提示共生菌群在内分泌介导的生长发育路径中发挥重要作用。
植物乳杆菌可恢复宿主正常发育、促进IGF-1分泌及活性,提高外周组织对GH的敏感性,内在的分子机制仍不清楚。
动物实验表明, 短链脂肪酸可以恢复动物的骨量、生长发育和 IGF-1水平,共生菌群的发酵产物(SCFA)可能在维持生命早期内分泌轴稳定和生长表型发挥重要调节作用,也暗示着共生菌群对婴儿发育迟缓和消瘦有影响。
营养不良及相关后遗症是一代一代相传的。
身材矮小母亲其子代的发育迟缓风险增高,代际传递在很大程度上归因于表观遗传修饰,这些修饰损害了后代生长发育。
近期两项研究提示:“营养不良”共生菌群的代际传递也会导致往后几代人的生长发育障碍,缺乏膳食纤维造成的共生菌群紊乱可在一代人的时间内获得逆转,但如果后代的饮食中再出现膳食纤维缺乏,则会在很大程度上不可逆转。
饮食引起的共生菌群菌种灭绝伴随着营养不良,是通过几代人营养不足的代际循环造成的。
5、生命早期以肠道菌群为靶点的营养不良疾病干预策略
肠道共生菌群结构和功能紊乱在儿童营养不良发生中起着因果关系的作用,有必要采用针对共生菌群为靶点制定策略预防并治疗营养不良。生命早期1000天是最佳的干预时间窗,共生菌群紊乱通过干预是可以治愈的。
妊娠期母亲与共生菌群互作影响着子宫内胎儿发育和出生后婴儿的成长轨迹,调节妊娠期母亲的肠道菌群可影响胎儿的发育。
例如,治疗产妇牙周病可降低低出生体重的风险;通过改善饮用水、环境和个人卫生可改善胎儿生长发育。
有关益生菌和益生元出生前干预的效果还不清楚,母亲孕期口服益生菌与早产率降低有关,但是最新一项荟萃分析并未发现孕妇口服益生菌或益生元对新生儿出生体重或其他方面有什么影响。
中低收入国家婴幼儿期暴露抗生素,促进婴儿产后持续生长,主要通过修饰肠道共生菌群来发挥作用。益生菌可改善高收入国家早产儿或低出生体重婴儿生长发育,有助于增加中低收入国家营养不良高风险儿童的体重。
最近一项4500例大样本临床干预试验发现,印度新生儿出生后2-4天内连续7天口服合生元制剂(植物乳杆菌+FOS)可显著降低新生儿60天内败血症发生率和死亡率,同时婴儿体重显著增加。
提示这种干预方案对败血症和营养不良高风险的早产儿或LBW婴儿获益将会更大,寡聚糖可被肠道共生菌群发酵利用,释放出SCFA和其它代谢物,这些物质有助于巩固婴儿小肠粘膜屏障。
粪菌移植(FMT)将健康受试者粪便完全置换患儿肠道菌群,有可能恢复那些传统治疗无效的急性重症营养不良(SMA)患儿的肠道功能和代谢稳态,促进生长发育,未来营养不良的疗法应考虑双重互补策略,结合营养素和共生菌群的干预策略,优化现有治疗方案。
6、未来亟待研究的问题
a) 营养不良患儿的肠道共生菌群结构和功能如何?
b) 营养不良和非营养不良患儿的母亲乳汁微生态结构有什么差异?它与儿童肠道菌群结构的关联如何?
c) 共生菌群在产前和产后子代生长发育的作用权重怎样?
d) 婴儿肠道共生菌群是如何导致婴儿发育障碍的?
e) 恢复共生菌群对急性重症营养不良接受营养康复治疗后的免疫功能影响如何?
f) 是否可以用共生菌群成熟度指数来预测发育迟缓或SAM的临床结局以确定高危患儿人群?
g) 在营养不足情况下,母亲阴道、肠道和口腔共生菌群在婴儿共生菌群定植建立中的作用是什么?这种代际传播是否会影响产后婴儿发育?
h) 益生菌能否有效地定植于营养不良患儿、发育迟缓的患儿肠道?
i) 哪一种共生菌群靶向的干预措施(抗生素、饮食、益生菌、FMT)能最有效持久地解决营养不良及其他相关临床预后?综合疗法是否更有效?
参考文献28篇(略)
(四)皮肤微生态与婴幼儿湿疹
皮肤是一个复杂的器官,由表皮、真皮和皮下组织三部分构成,其间还有丰富的血管、淋巴管、神经、肌肉以及由表皮衍生的皮肤附属器,后者包括毛发、皮脂腺、汗腺和指(趾)甲等。
皮肤从重量及面积角度看,是人体最大的器官,其总重量占体重的16%,表面积成人为1.2㎡~2.0㎡,新生儿约0.21㎡,新生儿和婴幼儿的体表面积要比儿童和成人的相对要大。
皮肤是人体的第一道保护屏障,其中居住着数量庞大的微生物(皮肤微生态),在抗感染、免疫平衡、屏障修复中均发挥着重要作用。
不同部位的皮肤,其皮肤微生态结构呈现较大差异。在皮脂腺分泌较多的(sebaceous)区域如脸颊,亲脂性微生物如丙酸杆菌属更容易富集。在潮湿(moist)部位如肘窝,则主要的细菌组分为葡萄球菌属和棒状杆菌属。足部的细菌组分主要也以葡萄球菌和棒状杆菌属为主。
除了细菌组分,真菌分布也有部位的差异性,如足部的真菌多样性更高,除了其他皮肤部位常见的马拉色菌属(Malassezia spp.),还包含黄曲霉菌属(Aspergillus spp.)、隐球菌属(Cryptococcus spp.)、红酵母(Rhodotorula spp.)、附球菌属(Epicoccum spp.)。
对比细菌和真菌,病毒的组成在部位间差异较小,而不同个体间的差异较大。
在上述提到的4种典型皮肤区域(sebaceous, dry, moist, foot),皮肤微生态的发展和稳定性也存在一定差异。通过随访1-2年,共3个时间点,对全身17个部位的皮肤样本进行宏基因组测序分析,总体显示个体的皮肤微生态随时空变化性较小。
但对比起来,足部和潮湿部位的皮肤微生态更复杂,同时稳定性也偏低,而真核病毒是导致足部微生态最不稳定的组成之一。肠道微生态主要受饮食结构等外部环境改变而发展,皮肤微生态的发展则受生理发育影响较大。
对比青春期前后的皮肤微生态发现:青春期会刺激皮脂腺产生更多的皮脂,从而使亲脂性微生物丰度增加,如:丙酸杆菌属(Propionibacterium spp.)和棒状杆菌属(Corynebacterium spp.),而发育之前的皮肤微生态则主要为链球菌属。
同时,发育之前皮肤的真菌多样性较高,发育之后则以单一的真菌马拉色菌属(Malassezia spp.)为主。
近年来研究也不断证实皮肤微生态和皮肤乃至系统免疫的相互影响。
不同于肠道微生态,皮肤上定植的微生物不参与真皮和表皮中免疫细胞的成熟,但可调节固有免疫因子的表达,包括:抗菌肽(如:cathelicidins和βdefensins),IL-1 alpha等。
然后,抗菌肽、补体系统和IL-1均代表研究较早的固有免疫系统,可能引起后续的宿主-微生物互作。同时,皮肤微生态相关的固有免疫,进一步引起系列适应性免疫反应,IL-1引导T细胞产生IL-17和干扰素γ。
在皮肤微生态和免疫系统互作中,免疫系统同样对皮肤微生态起着调节控制作用。近期研究证实,CD4+ 和CD8+ T细胞能够对接种于没有皮损的小鼠皮肤产生免疫反应,但该反应并不诱导触发炎症,皮肤免疫系统对皮肤定植微生物的这种免疫适应性通常被称为“动态平衡免疫”(homeostatic immunity)。
在碰到微生物入侵或感染时,皮肤免疫系统能够通过共生微生物的互作,产生一系列免疫反应,如T细胞能够促使角质层细胞分泌抗菌肽,从而阻止甚至清除病原体。
湿疹是国内外儿童期高发疾病,并可引起后续过敏性哮喘、食物过敏和过敏性鼻炎发病风险升高。前期研究发现皮肤微生态、免疫系统紊乱和湿疹发病以及疾病严重程度高度相关。
湿疹病人皮肤pH较高,同时皮肤微生态多样性显著降低。进一步研究显示:湿疹病人的皮肤微生态紊乱可以分为金黄色葡萄球菌为主,以及非金黄色葡萄球菌为主。
国外针对51个富集金黄色葡萄球菌的湿疹病人,45个未富集金黄色葡萄球菌病人和46个健康人的比较分析,结果显示金黄色葡萄球菌为主病人湿疹病情更重,II型免疫标志物(中性粒细胞数、总IgE量、CCL17、骨膜蛋白periostin)表达量更高,同时皮肤屏障紊乱更严重(经皮层水份流失TEWL高和角质层完整性低)。
其他研究表明:对比非金黄色葡萄球菌为主的病人,金黄色葡萄球菌为主的皮肤微生态多样性更低且湿疹病情更严重。
利用宏基因组学技术开展的一项研究进一步显示:皮肤微生态多样性和湿疹严重程度成反比,而金黄色葡萄球菌含量和病情成正比,同时病情较重的病人皮肤中金黄色葡萄球菌的基因组多样性较低,而较轻病人皮肤中的表皮葡萄球菌的基因组多样性较高。
而Donald Y M Leung 发表于2018年的研究显示:在富集金黄色葡萄球菌的湿疹病人中,富集MRSA的病人比富集MSSA的病人湿疹病情更严重,皮肤微生态的多样性也更低。
由于发育前后皮肤微生态结构的明显变化,青春期之前的湿疹患者(2-12岁)和青春期(13-17岁)及成年人(18-62)患者的皮肤微生态紊乱具有一定差异,青春期之前患者的皮肤微生态多样性更高,同时患湿疹后降低的主要是链球菌属,而青春期和成人患者降低的主要是丙酸杆菌。
2017年的随访研究揭露出生后第二个月肘窝有葡萄球菌富集,12个月时湿疹发生风险降低,提示早期皮肤微生态发展和湿疹发病的相关性。
鉴于皮肤微生态和健康的相关性,以及湿疹中皮肤微生态紊乱和宿主反应,近年来干预皮肤微生态预防或治疗湿疹成为关注热点。
研究较多的潜在皮肤有益菌是凝固酶阴性葡萄球菌(CoNS, Coagulase-negative staphylococci),主要包括:表皮葡萄球菌、人葡萄球菌和路邓葡萄球菌。这些潜在有益菌干预皮肤微生态的机制之一,就是直接抑制金黄色葡萄球菌的定植和感染。
近年来发表在Nature,Science Translational Medicine,Journal of Investigative Dermatology, Science Advance等杂志的研究证实:CoNS能够产生几种新型抗生素分子,从而有效清除金黄色葡萄球菌。
进一步的机制研究则表明皮肤共生菌能够阻断金黄色葡萄球菌群体感应系统,从而阻止金黄色葡萄球菌的进一步定植。
同时越来越多的动物模型实验结果显示:皮肤共生菌,尤其是CoNS,能够阻止金黄色葡萄球菌去激活角蛋白形成细胞中的丝氨酸蛋白酶活性,从而保护皮肤屏障的完整性,避免进一步的继发感染和炎症刺激,部分表皮葡萄球菌菌株更是被证实能调节免疫系统和引起组织修复。
除了动物模型研究,Richard L Gallo教授团队还尝试将人葡萄球菌应用于湿疹患者的皮损部位,结果显示24小时之内对金黄色葡萄球菌的清除率达到99.1%。
由于皮肤表面的生存环境中,微生物能够利用的营养缺乏,从而选择性利用特定微生物营养元素,如益生元,能够定向培养对皮肤健康有益的皮肤共生菌,从而抑制有害菌的定植并调节皮肤免疫的平衡。
表皮葡萄球菌能够利用甘油或蔗糖发酵产生特定结构的短链脂肪酸,若将表葡、丙酸痤疮杆菌和蔗糖共同注射到小鼠耳部皮肤,则巨噬细胞-炎症性蛋白2 (macrophage-inflammatory protein-2)和丙酸痤疮杆菌的含量均显著降低。
皮肤是人体的第一道保护屏障,早期研究逐步证实皮肤微生态的重要性,以及以皮肤微生态为干预靶标或有益菌资源库的临床应用前景。然而皮肤微生态和湿疹的相关性研究还面临系列挑战,也是将来需要重点关注的研究方向。
首先,需要明确目前的抗菌剂或系统性治疗方案对皮肤微生态影响的机制。其次,湿疹治疗中短期抗生素使用如何影响微生态,哪种抗生素对微生态影响更有害?
另外,皮肤微生态如何受局部用药、传统的非特异性免疫调节剂以及新型的目标疗法(如:Dupilumab,作用机制为阻断2型变态免疫性炎症的核心驱动因子IL-4/IL-13通路)的影响?
最后则是如何更精准的通过调节皮肤微生态,达到预防和治疗湿疹的临床目的。
参考文献27篇(略)
(五)呼吸道微生态与上呼吸道感染、 哮喘
呼吸道负责人体与外界环境的气体交换,主要由上、下呼吸道组成,前期研究已经证实呼吸道定植大量微生物且和呼吸道感染、免疫等相关。
研究较多的上呼吸道微生态为鼻腔、鼻咽和口咽3个部位,其中鼻腔和鼻咽微生态结构非常相似,主要由葡萄球菌属、棒状杆菌属、狡诈菌属、莫拉菌属、链球菌属等组成。
对比鼻腔和鼻咽,口咽部营养更丰富且更封闭,微生态组成结构也有较大差异,主要由链球菌属、普氏菌属、奈瑟氏菌属、韦荣菌属和嗜血杆菌属组成。
对比中国和欧洲、美国、澳大利亚的研究结果显示:不同区域健康人群的上呼吸道微生态结构相似,且环境影响大于宿主本身的影响。
由于声门的阻挡作用,初期认为在声门以下的下呼吸道和肺是无菌的,但随着支气管镜检查获取下呼吸道标本如支气管肺泡灌洗液(bronchial alveolar lavage fluid,BALF)技术的普及和不依赖培养组学技术的应用,目前发现在正常健康人的下呼吸道也有微生物定植。
国外的两项研究共纳入34个健康成年人,通过比较上呼吸道和下呼吸道微生态组成发现:下呼吸道微生态多样性比上呼吸道低,同时和上呼吸道相比,没有特殊的微生物组成。
肠道微生态在1岁以后趋于成熟,但多项队列研究显示上呼吸道微生态在出生后3-6个月趋于稳定。Bosch等采用16S rDNA扩增子技术分析102名健康儿童(从新生儿跟踪到6个月)的鼻咽样本。
结果显示出生后第1天,剖腹产和顺产新生儿的鼻咽均定植以链球菌属为主的微生态结构。出生1周内,鼻咽微生态迅速发展成葡萄球菌属为主的微生态结构,此后6个月葡萄球菌属逐渐减少,逐渐形成以棒状杆菌属、莫拉菌属、葡萄球菌属、链球菌属、狡诈球菌属和嗜血杆菌属为主的不同菌型。
另一项研究通过16S rDNA技术分析438名婴儿出生后1周、1个月和3个月的鼻咽样本,结果显示婴儿上呼吸道菌群可分为5种类型,前4种分别以葡萄球菌属、链球菌属、莫拉菌属、棒状杆菌属为主,第5种是混合菌属结构。
从出生后第1周到3个月上呼吸道微生态结构一直演变,如第一周时葡萄球菌属为主的结构最多,随着发育进程,莫拉菌属和棒状杆菌属为主的结构则逐渐增高。
欧洲的一项研究针对60个婴幼儿,分别取1.5个月、6个月、12个月和24个月鼻咽样本进行16S rDNA测序,结果显示:在1.5月龄时,鼻咽微生态结构已经形成,分别是以链球菌属、莫拉菌属、葡萄球菌属、棒状杆菌属以及棒状杆菌属和(或)狡诈球菌属为主的微生态结构。
对比鼻咽,口咽部可能受到饮食变化的影响,微生态结构稳定的时间稍晚,推测在1岁左右。
同时已经发表成果的队列研究显示:出生方式、喂养方式、抗生素暴露、有无兄弟姐妹、季节变化和疫苗接种对上呼吸道微生态产生一定影响,但进一步研究证实该影响是暂时的,和后续上呼吸道微生态发展和成熟的关联性,有待研究确认。
呼吸道感染是儿童期高发疾病,其中的下呼吸道感染如肺炎也是全球儿童死亡的主要原因之一。
无论是病毒还是支原体、细菌性病原体,感染后上、下呼吸道微生态多样性均降低,但不同病原体感染导致的呼吸道微生态结构的紊乱模式虽有差异,但鼻咽部均伴随着棒状杆菌属和狡诈菌属丰度的降低。同时,上呼吸道微生态的结构和感染后病情轻重高度相关。
美国一项多中心研究纳入1005名因合胞病毒感染入院的儿童,16S rDNA分析结果显示:病人鼻咽微生态具有4-5种结构,若以嗜血杆菌为主,则住院时长和进入PICU的比例高,而莫拉菌为主的结构则病情最轻,链球菌为主或混合结构的鼻咽微生态,则病情介于两者之间。
进一步研究显示鼻咽微生态结构与病情关系和宿主反应有相关性,如只有在抗菌肽LL-37≤46 ng/ml或CCL5≤42.0 pg/ml时,上述规律才明显。
而针对同一批病人的宏基因组和代谢组学分析显示:103个代谢物和临床是否使用positive pressure ventilation(PPV)高度相关,其中鞘磷脂代谢相关的通路和基因最明显。
进一步分析显示用PPV的患儿,微生物功能主要富集于链球菌属、罗思氏菌属、克雷伯菌属和嗜血杆菌属,而未用PPV治疗的患儿则主要富集于卡他莫拉菌。
上述研究均只能证实感染导致呼吸道微生态紊乱,但几项队列研究提示:上呼吸道微生态发展路径不同,急性呼吸道感染风险有差异。
针对澳大利亚234名新生儿,最终纳入177名婴儿在2、6、12个月时的鼻咽样本的随访研究,16S rDNA分析结果显示:2个月时嗜血杆菌属和莫拉菌属为主的鼻咽微生态,预示着1岁以内的急性呼吸道感染风险高,进一步分析显示春夏发病以嗜血杆菌属为主结构,秋冬发病则以莫拉菌属为主结构。
荷兰的112名新生儿随访项目中,采集了11个时间点的样本(出生后2小时、24小时、7天、14天、1,2,3,4,6,9,12个月),分析结果显示大部分儿童的鼻咽微生态发展轨迹为葡萄球菌为主---莫拉菌属为主---棒状杆菌和狡诈菌属为主。
然后对于一年年急性呼吸道感染频率较高的儿童,生命早期鼻咽部棒状杆菌和狡诈菌属丰度较低,同时口咽定植菌如奈瑟氏菌属、普氏菌属较多。
2019年针对芬兰的839个婴幼儿,通过16S rDNA技术分析2个月的鼻腔微生态,同时记录2-24个月期间的急性呼吸道感染发生率,结果显示莫拉菌属为主的结构,急性呼吸道感染发生率最高,而棒状杆菌属为主的结构发生率最低。
系列队列研究表明:生命早期鼻腔或鼻咽微生态紊乱,将提升上呼吸道急性感染风险,为此哈佛大学的Camargo教授提出上呼吸道微生态的“risk and resilience模型”,意味着不同的感染风险。
除了健康人群呼吸道微生态结构和发展,以及呼吸道感染和微生态的相关性,近年来的相关研究还涉及到呼吸道病原体的鉴定。
微生态分析技术能够鉴定呼吸道所有的病毒、细菌甚至真菌DNA或RNA片段,相关研究也证实该方法在临床上的价值,但目前仍有一定局限性。首先,呼吸道中本就定植大量微生物,甚至典型的病原体,如何有效区别定植和感染,将是后续研究的重点。
其次,如何更准确的对鉴定到的组分进行定量,并评估其毒力和耐药性也将是挑战。而技术本身的不足,如样本采集方法、评估污染、DNA或RNA的提取和富集、测序数据的分析、价格和周期等都将影响该方法在临床上的应用。
利用微生态分析技术进行临床病原体诊断之外,近年热议的研究方向还包括:利用呼吸道微生物之间以及微生物-宿主的互作,干预呼吸道微生态,从而预防或治疗相关疾病,如:慢性鼻窦炎、中耳炎、过敏性哮喘等。
近期发表的相关论文制备的鼻腔喷雾剂主要基于已有的益生菌(乳酸杆菌或双歧杆菌),结果显示使用鼻腔益生菌喷雾后,局部和系统免疫反应强于口服,而2017年针对20位慢性鼻窦炎患者的临床实验显示:虽没有任何副作用,但对疾病的缓解作用不明显。
后续呼吸道微生态干预的研究方向应在于:了解上呼吸道微生态的稳态结构中的“keystone species”,并评估其维持稳定中和宿主的互作关系和具体成分,从而调配合适的喷雾进行呼吸道微生态的干预。
参考文献18篇(略)
二、以宿主-菌群互作应答为靶点的干预策略
食物是我们生存和福祉的基本需求。饮食不仅对维持人类的生长、繁衍和健康至关重要, 而且还调节和维持着定居在我们人体消化道的肠道菌群。
得益于二代测序技术的方法学突破,在过去十年里,肠道共生菌群研究发生了显著的飞跃,揭示了不同微生物群落之间互作关系、集合功能和对宿主病理生理的影响,提供了健康和疾病期间肠道菌群变化的详细描述,以及对不同饮食方案的应答;
肠道菌群除了影响宿主生物过程的构建外,其组成和功能也具有可塑性,从而成为临床实践中治疗或干预疾病的一个有吸引力的靶点目标。
本文重点回顾以宿主-肠道菌群互作为靶点的几种干预措施的研究进展,膳食成分如何影响肠道生态、宿主生理和健康,现有研究证据给临床的启示,面临主要的挑战和未来研究方向。
(一)饮食
1、膳食纤维
膳食纤维是指任何来源、聚合度不少于3(DP≥3)的、不被人体消化吸收的碳水化合物多聚体、具有宿主生理健康益处的营养素;膳食纤维按以下几个参数进行分类,包括最初的食物来源、化学结构、水溶性、黏度、可发酵性。
大多数不溶于水的膳食纤维(如纤维素、半纤维素)和不降解的膳食纤维(瓜尔胶、圆苞)都有粪便膨胀作用,这是因为它们到达结肠后不被结肠菌群消化降解;
可溶性纤维不会引起粪便膨胀,而是被肠道细菌发酵,释放代谢产物如短链脂肪酸(SCFA);
一些高分子量的可溶性非淀粉多糖,如瓜尔胶、某些果胶、β-葡聚糖和圆苞车前子壳都是高黏性的,这意味着它们能够在肠道中形成凝胶结构,从而延缓脂类和葡萄糖吸收,影响餐后糖脂代谢。
并不是所有类型的纤维都存在于相同的食物里,抗性淀粉主要存在于谷物、豆类、块茎和不成熟水果像绿色香蕉等,而果胶则在水果和某些蔬菜中富集,谷类则含有β-葡聚糖和阿拉伯聚糖[3],由此推荐人类日常饮食应该丰富化和多样化。
膳食纤维贡献了肠道菌群的菌种多样性。
Yanomami土著人是迄今为止肠道菌群多样性最高的非工业化人群,其它非工业化人群(如巴布亚新几内亚农村人)的肠道菌群多样性也高于西方人群。
膳食纤维对宿主的影响表现在:
① 促进肠黏液分泌,降低肠腔氧气,维持正常肠腔生理功能;而长期缺乏膳食纤维导致肠道菌群结构改变,粘液层严重变薄,增加致病菌感染的易感性和慢性炎症性疾病风险;
② 促进宿主免疫系统的成熟和发育;研究证实膳食纤维代谢物SCFA可促进Treg细胞分化,并以GPR43依赖方式诱导组蛋白-H3乙酰化,相反,高脂饮食诱导胸腺不成熟,降低胸腺细胞数量降低,诱导发育中T细胞凋亡;
③ 提高膳食纤维摄入量有助于控制长期体重增加,减少超重和肥胖儿童的脂肪含量;
④ 2型糖尿病患者肠道缺乏降解纤维菌群,摄入可溶性纤维(寡聚果糖和长链菊粉)可纠正肠道菌群紊乱,遏制体重增加、改善低水平炎症,改善血糖代谢;此外,健康志愿者摄入含β-葡聚糖的面包可改善血糖代谢。
减少膳食纤维摄入不仅降低肠道菌群多样性和SCFA水平,还驱动肠道菌群代谢转向利用对宿主不利的能量底物,尤其是内源性蛋白质和宿主上皮粘液。
让受试者摄入高蛋白、低碳水饮食不仅显著降低了SCFA总量和丁酸的量,还导致了氨基酸发酵释放的有害代谢物增加,包括支链脂肪酸、氨、胺、N-硝基化合物、酚类化合物 (包括对甲苯酚、硫化物、吲哚化合物和硫化氢),这些代谢物的细胞毒性和促炎症作用诱导了慢性炎症性疾病的发生发展。
低膳食纤维饮食导致繁衍三代后小鼠肠道多样性发生大幅减少,即使恢复正常膳食纤维饮食后,其肠道菌群多样性也无法恢复,提示长期减少膳食纤维摄入可能导致重要的功能菌种永久灭绝。
最近一项队列研究观察到,当人们膳食纤维摄入量减少到30g/日后的24小时内,肠道菌群发生了戏剧性快速重组,不出所料,膳食纤维摄入量的骤减极大地降低了纤维降解菌群的丰度,肠腔SCFA浓度下降。
所有种类的膳食纤维都会导致肠道菌群结构发生某些特定的变化,哪些肠道菌群的结构变化有利于宿主以及对应的功能?
还有待进一步研究。膳食纤维促进宿主健康的作用机制主要依赖于代谢产物(如SCFA)、调节生理变化(如降低pH值)、或保护粘膜屏障。
膳食纤维影响肠道菌群结构表现出以下几个特性:
① 目前观察到的对人体肠道菌群变化都仅限于少数的几种谱系;
② 诱导变化的幅度可能很大,受影响的特定菌种可能占粪便菌群扩增后获得总序列的30%以上,但是只有摄入的时间足够持久才会维持这样的变化;
③ 肠道菌群对膳食纤维的应答是高度个体化的,这种个体化的原因还无法解释,推测可能原因是该个体缺乏具有特异降解酶、能利用特定底物的关键菌种或菌株。
综上所述,膳食纤维被看作是保护肠道菌群的关键性成分,有助于调节宏量营养素和宿主生理功能;从食物中提取和纯化的新型膳食纤维,选择性改进或合成膳食纤维,以发挥疾病预防和治疗的潜力;
未来肠道菌群靶向的膳食纤维干预可能会成为保健和饮食规划的重要工具,但膳食纤维和个体化饮食的确凿证据还有待进一步确定,今后的研究应集中于此类疗法的长期影响和安全性,以及它们可能扩大到肥胖及其代谢并发症之外的健康状况,如:营养不良、膳食成分缺乏症、炎症状态、肿瘤疾病。
2、个体化营养干预
众多因素影响着宿主与肠道菌群之间,以及宿主和肠道菌群对饮食应答的错综复杂的互作关系,一种饮食方案不能适应所有人群,在设计饮食方案时应该实施个性化干预。
精准饮食始于确定一个或几个与肠道菌群有关的变量,这些变量影响或修饰着饮食干预的结果。
研究发现,肠道宏基因组丰度的降低与饮食诱导超重或肥胖个体的体重丢失和体重维持成反比;
基线肠道菌群可预测饮食干预后有应答的受试者肠道中某种特定菌群的丰度,食用大麦内核制成的面包后血糖代谢获得改善的健康受试者,其粪便的普雷沃杆菌属/拟杆菌属比值较干预前显著提高;
而摄入人工甜味剂后出现糖耐量损伤的受试者,其基线肠道菌群结构与无应答的受试者存在显著差异。
随着先进的大数据分析方法的出现,现在可以破译多元因素间的互作关系,并提出精准的干预方案;
例如,一项基于小鼠肠道定植10种细菌的模型研究显示,用4种已定义食物(蛋白质、脂肪、寡聚糖和单糖)干预后,超过50%的肠道菌种丰度变化归因为饮食;
同样,基于宿主特定粪便菌种而建立的简单模型可以准确地预测健康女性对胆碱缺乏饮食诱发脂肪肝的风险。
最近一项研究运用机器学习算法,该算法基于肠道菌群基线结构和功能特征而构建,将800例健康受试者的个人参数纳入饮食规划,研究发现,它可以精准预测摄入各种食物后的血糖应答值,精准度超过目前广泛使用的碳水化合物计数或热量计数的金标准模型。
此外,根据个人预测的餐后血糖应答值实施短期饮食干预,可以成功地维持健康个体的正常血糖,在经过一周干预后,根据肠道菌群结构变化调整个人饮食,由此定期重新评估个性化参数并相应地调整饮食方案。
总之,精准饮食干预应根据个体的年龄、性别、地域、代谢状态、肠道菌群结构和功能,以及饮食偏好等个性化参数而构建。
事实上,研究发现,健康人摄入面包后的血糖应答在更大程度上取决于个人的基线参数而不是所摄入的面包类型,“适合的食物才是健康的”,这一结论反驳了普遍的认知,即“健康是指所消费食物的固有特性,因此有些食物比其他食物更健康”。
(二)微生态制剂(益生元、益生菌)
摄入益生菌的目的是补充肠道有益细菌,促进宿主代谢活性。
截至目前,益生菌干预治疗在许多方面都存在争议,虽然前瞻性随机对照试验(RCT)和荟萃分析结果证实,益生菌对抗生素相关性腹泻、哮喘、湿疹和炎症性肠病有效,但是绝大多数益生菌的临床试验结果不具有可重复性;
此外,许多益生菌的研究发现都是相关关系,缺乏潜在机制学研究,而且都是基于动物模式实验或体外条件下完成的实验,人群临床试验非常有限,因此目前还没有一个益生菌被FDA或欧盟批准用于医疗目的。
最近研究发现,益生菌对宿主影响作用不一致,主要源于益生菌肠道定植模式的个体差异及宿主肠道菌群特征。用基线宿主状态和肠道菌群特征,可预测益生菌在宿主肠道粘膜定植的“抗定植”到“可定植”。
未来益生菌应用于临床疾病治疗还面临着巨大挑战,包括:
① 开发直接从肠粘膜采样的非侵入性方法、以及能够可靠地鉴定肠道不同区域微生物群的技术;
② 需要确定益生菌菌株在体内的活动机制,以及预测治疗后肠道微生态的变化。
③ 要针对不同临床场景和不同人群亚群,完成大样本、多中心、前瞻性随机安慰剂对照临床试验,生成高质量和结论性的临床数据。
益生元是指能够被宿主体内的菌群选择性利用并转化为有益于宿主健康的物质。
益生元通过改善肠道菌群结构和功能而使宿主获益,以菊粉、低聚果糖(FOS)、低聚半乳糖(GOS)为形式的可发酵膳食纤维得到了广泛研究,认为可提高所有年龄组以及不同健康状态人群的粪便肠道双歧杆菌属、乳酸杆菌属丰度(后者又与SCFA提高相关)。
然而,现有研究证据存在局限性,包括:研究受试者和方法学存在很大异质性,疗效不具有可重复性,仅少数功效可真正使人群获益,如免疫调节、促进代谢或预防肠道致病菌感染,对益生元的应答也存在个体差异,取决于该个体基线肠道菌群特征。
其它益生元已完成小鼠和人群验证,确定它们通过调节肠道菌群而使宿主获益。
例如,健康受试者摄入大麦、全麦和褐米(每天60g或两者混合)可提高粪便菌群多样性,增加厚壁菌门/拟杆菌门比值,提高Blautia菌属丰度、改善餐后血糖峰值以及降低外周血IL-6水平;
以蔬菜和果汁为基础的饮食减少健康受试者粪便厚壁菌门和变形菌门丰度,提高拟杆菌门和蓝藻菌目丰度,并诱导宏基因组功能改变,提示有益于宿主代谢活性。
墨西哥传统医学中使用的仙人掌成分诺帕(即中医的黄连素)被认为可调节肠道菌群结构,促进SCFA产生,改善大鼠代谢表型。
其它益生元还包括:低聚糖、共轭亚油酸和母乳神经鞘磷脂,可改善高脂饮食诱导的小鼠代谢恶化。
令人惊讶的是,一些常用药物也具有益生元功效(例如,抗糖尿病药物二甲双胍,提高高脂饮食诱导肥胖小鼠肠道里AKK菌丰度),可能与提高肠道杯细胞数量有关,也暗示通过调节肠道菌群发挥抗糖尿病作用。
值得注意的是,益生元的定义已经获得修订,强调其对与宿主生理学相关的微生态和功能特征的影响,而不是专注于选择性细菌的特定活性。
(三)粪菌移植(FMT)
粪菌移植(FMT)是人类最古老的基于肠道菌群的干预措施,可以追溯到第四世纪。
在一项具有里程碑意义的研究中,将健康人新鲜粪便粪菌液经十二指肠输注给复发性艰难梭菌感染患者,对照组采用标准的万古霉素治疗,随访10周后显示FMT组复发率显著降低。从此FMT开始用于其他疾病的研究,如代谢性疾病和IBD。
FMT能成功地治疗代谢性疾病,但对IBD的疗效还存在争议;
一项非随机研究选择6例溃疡性结肠炎患者经结肠镜实施FMT,经过多次治疗肠道菌群结构显著改善,但未达到临床缓解;
另一项对50例临床轻中度UC患者的随机对照试验显示,经鼻-十二脂肠镜实施FMT的临床缓解率为30%,自体FMT对照组为20%;
Moayyedi等随机选取75例轻中度UC患者,实施灌肠FMT,结果显示干预组应答率24%(22例),而对照组5%(仅1例)。
另一项研究对41例类固醇药物依赖的活动性UC患者先实施结肠镜输注FMT再实施灌肠FMT,干预组有27%达到无需固醇药物的临床缓解,而对照组仅8%。
最近完成的一项对41例固醇类药物依赖UC患者实施FMT观察发现,治疗后第24周,干预组无固醇类药物的临床缓解率为46.3%(19/41),临床应答率和镜下缓解率分别为75.6%(31/41)和63.4%(26/41),所有临床应答患者都停用了类固醇药物。
最近,FMT治疗IBD的荟萃分析收录了31项研究报告,其中亚洲受试者较西方受试者年龄偏小,其UC和CD的临床缓解率分别为39.6%和47.5%。
另外一项综述报告分析,捐赠者粪便是否新鲜或冷冻、FMT实施途径以及抗生素预处理都不会影响IBD疗效。
非酒精性脂肪肝(NAFLD)也是FMT的应用指征,目前FMT对NAFLD临床研究的疗效还未有结论,高脂饮食诱导大鼠脂肪性肝炎动物模型已经证实FMT疗效[40],干预组HFD小鼠接受FMT后肠道菌群结构显著改善,克里斯滕森菌科(Christensenellaceae)和乳酸杆菌科丰度上升,小鼠盲肠丁酸浓度显著提高,肠粘膜紧密连接蛋白ZO-1表达上调,镜下HFD诱导脂肪性肝炎显著改善,提示FMT对NAFLD有潜在治疗作用。
FMT的一个局限性是由于未知的因素,不同的粪便捐献者疗效不同。另一个问题是传播传染病或由肠道菌群介导的、捐献者传到给接受者的某种特质。
一种可替代的方案,也是更具个性化的方案是自体FMT,就是在发病前存入该个体粪便样本,再进行FMT,这种方案需要大规模的粪便银行设施,这个方案仍然有潜在风险,因为看起来健康的个体可能携带某种需要治疗的致病因素,导致不可预见的疾病发生。
未来的研究应该探索能够确保FMT捐赠者优于其它捐赠者的因素,破译移植的粪菌与宿主原生肠道菌群之间互作关系,并阐明肠道定植情况,新的机制学探索有助于开发涵盖不同功能的、肠道菌群定制的“设计师”治疗策略。
(四)菌群代谢物(后生素)
共生菌群靶向疗法的另一个例子就是使用细菌衍生代谢物,即后生素(Postbiotics)。
它是指由活菌分泌的代谢产物或细菌死亡溶解后释放的可溶性因子,这些物质通过提供额外生物活性有益于宿主的生理功能,包括但不限于:短链脂肪酸(SCFA)、酶、肽、磷壁质酸、muropeptides、胞内和胞外多糖、细胞表面蛋白、维生素、纤溶酶原和有机酸。
研究已经证实,肠道菌群广泛地影响着胃肠道和肠道外器官生理功能,包括:抗菌、抗氧化和免疫调节;
近年越来越多的研究支持这些获益主要依赖于菌株分泌的代谢物或衍生因子,这些化合物通过调节肠道菌群稳态和/或宿主代谢和信号通路,影响宿主特定的生理、免疫、神经激素生物、调节和代谢反应。
后生素可根据化学结构分为:脂类(如:丁酸、丙酸、二甲基乙酰衍生的纤溶酶原)、蛋白质(如:乳酸素和P40)、碳水化合物(如寡聚半乳糖和磷壁质酸)、维生素/辅酶(如B族维生素)、有机酸(如:丙酸、3-苯基乳酸)、复杂分子(如Muropeptides和磷脂酸);
或者根据功能分类,如调节免疫、抗炎症、降低胆固醇酯、抗肥胖、抗高血压、抗增殖和抗氧自由基作用。后生素具有多种令人瞩目的特性,如化学机构清晰、安全剂量明确、半衰期长(5年);
此外,后生素制剂有良好的吸收、代谢、分布和排泄特性,暗示在宿主体内不同器官和组织的信号能力很强,有助于引发多种生物学反应;后生素可模仿益生菌的健康效应,同时又避免了活菌制剂制备和通过胃肠道时的衰减。
大多数情况下,后生素源自乳酸杆菌和双歧杆菌的菌株,也有报告选用链球菌和粪杆菌菌株;研究证明,后生素制剂可降低血压,推测可能是改变了肠道菌群结构和代谢产物谱,重建肠屏障功能,改善内毒素血症,降低炎症水平和调节神交感神经活性。
酵母代谢也能产生后生素,布拉氏酵母菌培养后上清液通过体外激活α2β1整合蛋白受体,提高了伤口愈合能力和上皮细胞迁移,在各种肠道疾病中具有潜在治疗作用。
后生素促进宿主健康作用的详细分子机制尚未充分阐述,现有研究提示作用机制不尽相同,其中,后生素激活过氧化物酶体的增殖-激活受体,加速脂肪酸β氧化,降低甘油三酯;
也有研究发现后生素激活核苷酸结合寡聚蛋白1,诱导脂肪细胞自我降解,降低肝组织HMGCS和HMGCR酶活性,提高肝脏和肌肉组织的AMP激活的蛋白激酶,促进脂肪分解代谢,控制血脂异常。
此外,一种叫甲酰二肽的后生素通过核苷酸结合寡聚蛋白2和激活转录因子 IRF4路径改善肥胖小鼠脂肪组织炎症和葡萄糖不耐受。人体研究证实,后生素能改善肥胖和全身低水平炎症,提高肝组织胰岛素敏感性。
后生素还具有特异性抗结肠癌细胞增殖作用,抗癌机制与调节免疫应答,诱导细胞凋亡有关;
从乳酸杆菌菌株获取的后生素降低间质金属蛋白酶-9活性,抑制结肠癌浸润;进一步研究发现, 活性抑制评分与分子量 > 100 kDa 和 50–100 kDa 的化合物相对应,提示该抑制性化合物可能是蛋白质、核酸或多糖等大分。
表多糖也具有抗氧化作用,一种来自瑞士乳杆菌MB201的表多糖带有大量负离子,能够螯合尿酸上的亚铁离子,亚铁离子参与了自由基形成,从而产生活性羟基自由基。
后生素的抗菌活性归结为已知和未知的抗菌化合物,包括但不限于细菌素、酶、小分子和有机酸,对革兰氏阳性细菌和革兰氏阴性细菌表现出抑菌或杀菌特性。
后生素通过提供更好的和更具特异性的生理功效,有助于改善宿主健康状况,未来还需要进行更多的研究以阐明这些化合物的预期和非预期效应。
相比益生菌食品,后生素具有更稳定、安全性高、生产和应用中品质可控和可标准化,在功能性食品领域比益生菌有更广阔的应用前景。
未来需要做出更多的研究,发现和鉴定新的后生素产品;进一步研究了解后生素作用的信号通路,确保后生素产品制造过程中的稳定性及其功效;
在制定统一和严格界定的培养程序时,控制和消除后生素制造过程中的可能变异;此外,还需要设计良好的随机安慰剂对照的临床干预试验和代谢组学研究,审查人体长期的安全性,为产品的健康宣称提供医学证据。
(五)噬菌体
噬菌体是一类能够在细菌细胞中感染和复制的病毒。近一个世纪前曾首次使用噬菌体疗法,当时主要被用来治疗各种疾病,如:伤寒、痢疾、皮肤和手术伤口感染、腹膜炎、败血症、尿路感染和外耳炎。
1930年代一系列综述研究报道噬菌体在质量控制、标准化制备以及临床疗效不一致等方面颇受争议;自1940年人们的关注点聚焦在抗生素疗法上,噬菌体疗法的研究局限在东欧几个国家,语言限制了研究结果的传播。
直到新千年开始,全球性抗生素耐药危机驱动了噬菌体疗法的复苏,此外,我们对噬菌体生物学、遗传学、免疫学和药理学认识的提高也促进了人们对噬菌体疗法的新兴趣,对噬菌体治疗的关键方面已经标准化,显著提高了治疗成功率。
相比抗生素疗法,噬菌体疗法的最大优势是:
① 特异性高,不杀灭共生菌群也不亲和人体真核细胞,副作用比抗生素少很多;
② 自限性,一旦宿主细菌被杀,其杀菌功能就停止;
③ 在宿主细菌内部呈指数繁殖,临床治疗所需剂量较低;
④ 具有抗生物膜作用,噬菌体体外编码的解聚酶可降解生物膜基质,从而更好地促进抗生素和噬菌体颗粒渗透进致病菌菌体内部发挥杀菌作用。
目前噬菌体疗法已经进入美国临床Ⅰ/Ⅱ期试验,适应症和给药方式包括:局部给药治疗慢性静脉炎腿部溃疡(靶向致病菌:金黄色葡萄球菌, 铜绿假单胞菌, 和大肠杆菌)、局部治疗耐药性绿脓杆菌所致慢性中耳炎、口服治疗儿童腹泻和成人急性胃肠炎,上述研究一致性认为噬菌体疗法具有良好的安全性和耐受性。
临床上静脉或腔内噬菌体疗法主要通过知情同意使用的个案研究进行的,如美国首例用静脉注射和腔内噬菌体制剂成功治疗全身多器官衰竭性感染的病例;
一名76岁男子接受主动脉弓置换手术,术后确诊全身绿脓杆菌MDR感染,在抗生素治疗无效情况下采用噬菌体鸡尾酒疗法,结果显示受试者耐受性很好,绿脓杆菌感染在抗生素停用后没有复发;
一例61岁患者罹患绿脓杆菌性败血症,罹患急性肾损伤不耐受抗生素治疗,开始接受双噬菌体鸡尾酒疗法,连续治疗10天,噬菌体疗法后血培养转为阴性,C反应蛋白降低,患者体温恢复正常,数天后肾功能恢复。
Speck和Smithyman最新做了系统回顾,他们重点评价过去80年来利用噬菌体静脉给药对伤寒和金葡菌性菌血症等重症感染的疗效,收录了近1000例成功接受静脉噬菌体治疗的患者,副作用的报道数量微不足道。
所有报告严重的副作用(类似休克或血清疾病反应)都可归因于早期噬菌体制剂生产过程中带入的污染物,最后结论认为:噬菌体静脉给药在很大程度上是安全和有效的。
噬菌体疗法尽管潜力巨大,但同时也面临重大挑战,包括无法在体内模拟体外抗菌效果、无法体内重复同样杀菌效果,可能原因包括剂量问题、噬菌体诱变、与肠道菌群互作、中和宿主抗体、或出现噬菌体耐药致病菌株。
针对不同致病菌群的靶向受体的噬菌体组合能提供上述问题的部分解决方案。
参考文献33篇(略)
三、国内外几项出生队列研究简介
越来越多的研究提示一个惊人的关联:许多儿童和成人复杂性疾病,如出生缺陷、孤独症、多动症、哮喘、代谢综合征、糖尿病、不孕不育、肥胖、心脑血管疾病等都与生命早期环境暴露关系密切。
“发育起源学说(Developmental origins of health and disease, DOHaD paradigm)” 认为:
生命早期暴露与远期健康结局关系密切;而生命早期环境因素作用具有时效性、隐匿性和累积效应;由此,全球范围内开始建立出生队列研究,探讨孕前、孕期及出生早期环境暴露对婴儿、儿童、青少年、成年人及老年人健康的影响;
出生队列研究通常从母亲孕期或孕前期招募研究对象,采集父母双方基线信息、孕期各阶段暴露信息,以及各种类型的生物样本(包括:孕产妇静脉血、尿液、胎盘、脐带血、母乳、胎儿粪便、儿童尿液、静脉血、头发),进而对子代进行长期随访,定期搜集新生儿期、儿童期乃至成年期以后的各类暴露信息和生物样本,获取各类疾病或健康有关的结局变量。
这样的研究设计可以实现对人体生命周期中关键环节的关键暴露和关键结局进行准确的收集、整合与分析,获得暴露因子与疾病之间有无因果关联及关联性大小信息,是目前公认的疾病病因学研究的“金标准”;
以下简要回顾国外重要的几项出生队列研究,并简单介绍中国大陆的三项正在进行的出生队列研究。
(一)英国儿童发育研究(National Child Development Study, NCDS)
英国1958 年出生队列(1958 British birth cohort,NCDS)是一项持续性、多学科的纵向队列研究,始于1958年,2008年进行了最后一次的测量和评价。
最初是由英国国家生日信托基金会(National Birthday Trust Fund, NBTF)发起的围生期死亡率研究,调查1958年3月3-9日出生于英国的17415名儿童,旨在寻找与死产和新生儿死亡相关的社会学因素和产科因素,提高英国的产科服务,降低围生期死亡率;
随后由英国国家儿童局(National Children’s Bureau)负责跟踪调查对象7岁时的身体、教育和社会方面的发展情况。
目前已经进行了9次调查,分别在研究对象7、11、16、23、33、42、46、50和55岁时,通过问卷调查、医学检查、认知评估、面对面访谈等方式收集资料,最后一次调研是在2018年受试者60岁时完成。
收集的主要资料包括:
健康和医学资料(包括:免疫接种、体格发育、情感发育、疾病、生育情况、医学指标),人口统计学资料(家庭背景、社会经济条件、认知发育、受教育情况、就业、社会心理学特征、人际关系、健康相关行为);
目前主要资助来自于英国经济和社会研究理事会(Economic and Social Research Council),由英国纵向研究中心(Centre for Longitudinal Studies)承担调研资料收集。
主要研究发现:
1、母亲孕期吸烟造成子代近期和远期危害。
母亲孕期吸烟可导致子代围生期死亡率的升高和出生体重下降,它对子代身高发育影响较小,但是对子代的智力发育有很大影响。
母亲孕期吸烟不但增加子代从儿童期到成年期的肥胖发生风险,还引起成年中期向心性肥胖等心血管疾病的发生风险;母亲在孕期吸烟是子女成年早期罹患Ⅱ型糖尿病的危险因素,同时也显著增加子代儿童期呼吸道疾病发生率;胎儿期父亲吸烟的儿童其Ⅰ型糖尿病(T1D)的患病率较低。
2、家庭环境、事件对儿童近期和远期影响。
儿童期家庭不和谐导致7岁前生长发育缓慢,住房拥挤与7岁时身材矮小存在关联。暴露于不利因素如父母离婚、社会经济地位低的儿童在青春发动之前身高增长相对缓慢,在青春发动之后呈现出明显的追赶生长现象,但是,最终并不能达到非暴露者的水平。
父母分居或离婚对子代成年期的心理健康有影响,如成年早期情感障碍的发生率升高、学习成绩较差等, 成年期心理症状和酗酒的发生风险增高,但是心理症状风险的增高在成年早期( 23岁、33岁) 就已表现的很明显, 而酗酒风险的增高则在后期才开始显现。
另外,儿童期暴露于不利因素如经济困难、父母对其教育缺乏重视、暴力事件等的女性, 其子代的不良出生结局如低出生体重、早产增加。
3、儿童期疾病和发育障碍对青春期、成年期的影响。
儿童时期心理障碍对个人一生的影响远比身体健康问题重要得多,不仅成年期的工作和收入,还影响代际间及代内的社会阶层流动性,儿童期经历心理障碍者到50岁时家庭收入减少了28%。
儿童期轻度精神发育迟滞对成年期生活有影响,许多人表现出一种持续性的损伤,如成年期抑郁症的发病率升高,那些成年期患有精神疾病的人往往有儿童期精神分裂症、社会适应功能失常的病史。
儿童期问题行为对于从儿童期到成年期的终身伤害风险具有预测作用,外化性行为增加终身伤害风险,主要包括交通事故、家庭暴力、工作场所伤害以及暴力所导致伤害,而内化性行为减少终身伤害风险。
(二)丹麦国家出生队列(Danish National Birth Cohort, DNBC)
1996年启动的丹麦国家出生队列研究(DNBC)就通过收集生命早期(孕期和出生后几年)各种因素暴露,对各种复杂性疾病进行病因流行病学研究,特别肥胖症、代谢性紊乱、儿童哮喘和过敏症的发生原因。
丹麦等北欧国家一直以来就有全人口覆盖的个人疾病登记、人口统计学和社会经济状态的注册制度,通过个人ID号将所有居民个人信息关联起来,这项研究一开始就成为国家战略项目,获得丹麦国家卫生委员会下属国家研究基金会资助,丹麦三所大学和国家血清研究所负责实施,由全科医生和助产士负责收集生物样本。
该项队列研究的招募工作从1996年开始,到2003年结束,自母亲怀孕开始就接受定期随访跟踪,直到孩子满18岁结束,信息收集工作于2014年9月完成。
整个随访期间受试者接受4次电话采访(分别孕12周、孕30周、产后6个月和18个月),当孩子年满7岁、11岁和14岁时,家长收到一份关于孩子生活和生长发育的调查问卷,收集了关于儿童生活、生活方式、饮食习惯、健康和福祉的信息;
18岁后这些青少年通过邮件回答调查问卷,随访内容包括有关情感障碍、注意力缺陷多动障碍、饮食行为紊乱、自残和自杀的问题。
此外,队列研究建立了一个生物库,收集孕期母亲两次静脉血(孕12周、孕24周)、刚出生后不久采集脐带血。
这项研究共纳入101,042名孕妇、91,256名7岁儿童、43,265名18岁成人。截至2019年4月共计发表570篇研究文献(含11篇队列研究方法学研究报告)、45篇博士论文。2019年5月29日丹麦DNBC管理层宣布在哥本哈根召开科学研究会,以此纪念DNBC过去20年的研究亮点。
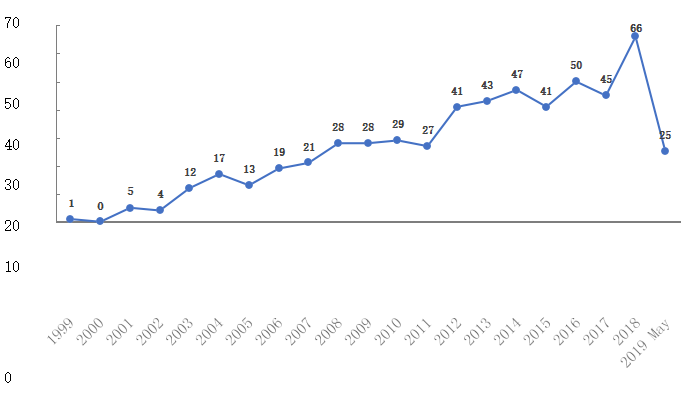
图2:丹麦出生队列研究1999-2019年发表的研究文献(559篇)
(三)荷兰KOALA出生队列(KOALA Birth Cohort Study)
KOALA是荷兰语“儿童、父母和健康:生活方式和遗传物质”的首字母缩写。这项出生队列研究是迄今为止首次大规模前瞻性观察肠道菌群与特应性疾病发生之间关系,并对多个混杂因素进行控制,已验证肠道菌群干预宿主免疫系统发育,参与特应性皮炎发生发展的研究假设。
研究主要关注生活方式与肠道菌群的建立和发育,如:父母饮食模式、疫苗接种、抗生素、喂养方式、肠道菌群结构、出生后第一年感染,以及基因与环境的相互作用。
本项研究招募工作从2000年开始,直到2012年结束;招募对象是孕妇和2001-2003年期间出生的新生儿,共计招募2834名孕妇,其中2343名孕妇属于“传统生活方式”,从正在进行的与怀孕相关的盆腔腰带疼痛前瞻性队列研究中检索出来的;
另外491名孕妇属于“替代生活方式”,通过有机食品商店、嗜食症诊所或助产士招募而来,有助于观察各种生活方式(如:家庭育儿、有机食物、素食饮食、限制或不接受疫苗和/或抗生素)对儿童发育和疾病发生的作用。
所有受试者都在怀孕14至18周之间登记建档, 同时完成一份关于特应性皮炎家族史和婴儿护理意向的录取问卷。
在怀孕期间和婴儿出生后头两年内第3、7、12和24个月里, 所有受试对象接受重复调研问卷确定关键性暴露因素,调查问卷收集了有关卫生、感染、营养、育儿和其他生活方式特征的信息。
同时,该研究还采集了生物样本, 包括:孕母外周血(孕期36周时采集以确定IgE水平)、婴儿粪便(产后1个月采集,分析婴儿肠道微生物群)、母乳(在产后1个月采集,确定母乳脂肪酸和免疫标志物水平)、婴儿1年和2年后静脉血(测定感染后血清学指标和IgE水平)。
所有婴儿及其父母都用Bucal试子擦拭收集脱落细胞,提取用于基因分析的DNA。
为确定婴儿过敏症诊断,在婴儿7、12、24月龄时由家长完成儿童哮喘和过敏症的国际调研问卷,同时婴儿2岁时护士进行家访观察过敏性皮炎情况。
主要研究结果:
目前该队列研究已累积发表研究文献56篇,其中最具创新性的发现是:定居在肠道内的共生菌群是新生儿出生后最关键重要的暴露。
KOALA队列中相关肠道菌群的研究结果为健康婴儿肠道菌群发育提供了以下关键性见解:
①出生方式对健康婴儿肠道菌群的建立和成熟至关重要,足月、阴道产和在家分娩有助于肠道有益菌群的定植和繁殖;
②婴儿饮食显著影响着健康婴儿肠道菌群器官的发育,绝对母乳喂养有利于肠道有益菌群繁殖;
③肠道菌群与过敏症:婴儿肠道里某种特定菌群与湿疹、复发性喘息和过敏症高度相关。
1、影响早期肠道微生物群组成的因素
本研究采用PCR分析技术对1032例1月龄婴儿的粪便样本进行了特定菌群分析,包括双歧杆菌属、乳酸杆菌属、埃希氏大肠杆菌、艰难梭菌、拟杆菌属、微生物总量。
采用单变量和多变量分析来确定各种菌群与产妇问卷的多种因素的相关性;研究发现,相比阴道分娩的婴儿,剖宫产婴儿肠道缺乏双歧杆菌和拟杆菌属,而富集难辨梭菌;绝对奶粉喂养婴儿肠道富集埃希氏大肠杆菌和难辨梭菌,而母乳喂养婴儿肠道富集双歧杆菌和拟杆菌。
早产儿和住院婴儿肠道富集艰难梭菌;服用抗生素的婴儿肠道双歧杆菌和拟杆菌丰度降低;家里有兄弟姐妹的婴儿肠道双歧杆菌丰度显著高于没有兄弟姐妹的婴儿。研究结果显示,足月、阴道产和家中出生的婴儿肠道有益菌群数量最高。
2、肠道微生物群在特应性疾病发生中的作用
采用PCR技术检测957例婴儿粪便样本,婴儿2周岁时采集血液样本测定IgE总和和特异性IgE水平,并通过产妇问卷收集有关特应性症状的信息,婴儿2周岁时对特应性体征进行临床诊断。
研究表明,肠道埃希氏大肠杆菌丰度增高的婴儿罹患湿疹的风险增加,并具有量效关系;肠道富集难辨梭菌的婴儿罹患湿疹、复发性喘息和过敏症的风险增高;肠道检出难辨梭菌则显著增加特应性皮炎发生风险。
更重要的是,研究表明,肠道菌群结构先于特应性皮炎的发生,而埃希氏大肠杆菌和难辨梭菌与皮炎发生相关。
(四)挪威母婴队列研究(The Norwegian Mother and Child Cohort Study, MoBa)
挪威MoBa研究是一项从怀孕开始到子代出生并长大至19岁的纵向观察性研究,该项目自1998年获得国家议会投票通过后,于1999年正式启动,由卫生部和国家卫生研究所提供种子资金(1999-2008年约为4500万美元),挪威公共卫生研究所 (NIPH)负责项目实施。
本项研究的目的是通过验证特定病原假设,预测暴露因素(包括遗传因素)与目标疾病之间的因果关系,为制定预防策略提供依据。
由于人们无法预测未来10-50年出现的具体问题,MoBa规划方案并不是根据任何单一的假设,而是收集尽可能多的相关暴露和健康结果的数据。
这项研究计划以家庭(父亲+母亲+孩子)为核心、以单次怀孕为基本单位,招募在1999年到2008年期间所有受孕并分娩的妇女,从怀孕第17周实施超声扫描并建立健康档案开始,全部孕期共随访三次(孕17、22、30周),子代出生后开始跟踪随访至15岁,共随访九次(生产后、6月、18月、36月、5岁、7岁、8岁、13岁、14岁);
1999-2005年期间实际招募了64,136名孕妇,占这个期间全部孕妇45.0%;截至2016年底,该研究已经招募113,890名孕妇、113,490名儿童(其中双胞胎1950对,三胞胎21对),2.7万个家庭,包括孕妇及其孩子和丈夫共计27.0万人。
收集的资料信息包括:
① 父母和家庭基线信息;
② 各种类型生物样本(母亲尿液、血液;父亲血液;子代脐带血、尿液、外周血);
③ 健康调研问卷(孕期母亲3次,父亲1次,产后6个月、18个月、3岁、7岁时各1次),第一份调查问卷是在怀孕第13-17周进行,收集母亲怀孕前和怀孕期间的病史、药物、职业、工作场所和怀孕期间的病史的数据,以及家庭、生活习惯和心理健康信息;
第二份调查问卷在孕22周时发出食品频率调查问卷;
第三份问卷在孕3周时发,信息涵盖妇女在怀孕期间的健康状况以及工作情况和习惯的变化;
第四份问卷在子代出生后6个月,重点收集儿童健康和营养以及产妇疾病、福祉和心理健康资料;
18个月、3岁和7岁的调研问卷主要关注儿童的生长发展、身心健康和疾病状况;
④ 国家卫生登记信息(医院和门诊疾病登记、癌症登记、处方数据库、死因登记和疫苗接种登记)。
MoBa研究的优势是在一个研究中可同时测量基因和环境因素,尤其是孕妇队列具有特别价值,收集了孕期的环境暴露;生物标志物可反映疾病发生发展的早期和亚临床迹象;
本研究项目截至目前发表研究文献460多篇,共产生超过220项子设计,与疾病病因探索相关的子设计包括:自闭症(Autisum Birth Cohort,ABC)、儿童多动症(ADHD study)、儿童IBD、哮喘、母亲和子代癫痫、婴幼儿神经系统和语言发育、儿童糖尿病;
尤其值得一提的是,MoBa队列数据参加欧盟资助的国际合作研究(欧洲、亚洲、南美洲):先兆子痫的全基因组关联研究 (GWAS),研究发现胎儿基因组中临近编码FLT1(FMS样酪氨酸激酶1)的一系列基因位点发生变异提高了先兆子痫发生风险,这个新发现的基因位点提高了人们对先兆子痫及各亚型发病机理的理解。
(五)芬兰出生队列研究(The FinnBrain Birth Cohort Study, FinnBrain)
芬兰出生队列研究(FinnBrain)是一项研究探索生命早期生活压力对儿童神经心理发育影响的纵向队列研究,其目的在于通过前瞻性、多学科队列研究,探讨产前和生命早期生活压力对儿童大脑发育的影响,根据研究发现的新靶点实施干预研究,为促进儿童身心健康发育,预防不良结局干预策略的制定提供依据。
该项队列研究包括几个子项目,其中三个重要的子课题为:
① 早期生活压力与儿童神经心理 (压力和情绪调节、气质、执行功能)的发育;
② 早期生活压力与“脑-肠轴”发育;
③ 早期生活压力与儿科传染性疾病。
队列招募工作是在该国家三个产妇福利诊所,招募对象是孕期连续进行免费超声波检查的妇女,重点是父母群体(包括妊娠和第一次产后婴儿评估),招募自2011年12月开始,至2015年4月结束,共纳入3808名母亲、2623名父亲/配偶、3837名儿童(含29对双生子),共计5790个家庭。
数据信息获取方式:
① 调研问卷,自妊娠3个月开始入组,母亲在怀孕期间和分娩后分别三次填写调研问卷(孕14、24、34周和分娩后0-3天),生产后至子代3-4岁期间母亲和子代分别连续8次填写调研问卷(产后1-2.5月、3、6、8、12-24、18-24、30、36-48个月);
调研问卷重点收集反映个人和家庭的产前焦虑症(PS)和生命早期压力状态和自我修复能力,系统评价家庭生活事件、日常麻烦、生活和关系满意度、身体健康、抑郁和焦虑症状、睡眠和药物使用情况;
此外,父母调研问卷还收集反映子女养育的特征,如:父母的童年经历和与自己父母的关系、父母自己的依恋模式、父母自我反思能力和养育压力;儿童问卷的重点是气质和情绪调节、心理和语言发展、身体健康 (如感染、特应疾病) 和睡眠等方面;
② 国家登记注册数据(体检、住院和疾病医院登记、处方药数据库、死因登记和疫苗接种登记);
③ 各类生物样本(血液、尿液、粪便、母乳、子代头发、胎盘);
④ 仪器设备检查(大脑MRI、超声波、皮质醇);
⑤ 身心状态评估量表(爱丁堡产后抑郁量表、整体焦虑症状量表、怀孕相关焦虑问卷)。
主要研究发现:
父母教育水平较低与抑郁、焦虑和妊娠相关焦虑症状升高有关,这种相关性贯穿全部妊娠期,妇女和男子的情况都是一致的;而且这种相关联系在产后早期女性仍然明显, 但在男子则不明显。
母亲孕期焦虑提高婴儿6个月时负面情绪应答,支持产前因素在儿童情绪调节发展中的重要性,强调妊娠忧虑时早期压力事件的来源之一。婴儿的气质是影响早期母子关系的重要因素,婴儿的负面情绪降低母婴粘合关系;母亲的兴趣轨迹可预测婴儿的兴趣调节和气质特征。
孕妇产前焦虑和抑郁症状与一组“促过敏性”细胞因子和白介素(IL-5, IL-9和IL-13)水平正相关,为进一步研究孕妇产前状态与儿童健康之间的关联性提供了假设。
产妇的产前压力提高了婴儿期恐惧反应,产妇产前压力和母乳皮质醇浓度都与女婴的恐惧反应高度相关;产妇产后焦虑预测女孩的执行力较差,而产前焦虑预测女孩和男孩执行力存在性别差异。
研究表明,不同形式的早期生活压力( ELS)影响婴儿反应和自我调节的各个方面,这些影响存在性别差异。
(六)日本21世纪新生儿队列(The Longitudinal Survey of Newborns in the 21st Century)
日本2001年启动的“21世纪新生儿队列研究”目的是解决低出生率和人口老龄化,以及相关社会保障等问题,全部研究资金由日本卫生、劳动和福利部 (MHLW)提供资助,这项队列研究是世界上为数不多的全国性出生队列研究之一。
调查的目标人群是2001年1月10-17日以及7月10-17日出生的、并在日本居住6个月的所有新生儿,共计53,712名婴儿,通过邮件发给这些婴儿家庭,收到47,015份回答完整的调查问卷,覆盖率达87.8%。
该项目每年定期通过邮件向参与者发送调查问卷,前10次调查均有监护人填写完成,第11和12次调查(此时研究对象进入小学高年级)的管理方式不同,问卷前半部分由监护人完成,下半部分由儿童自行完成;
第13和14次(研究对象升入初中)则分别进行两份独立问卷调查,一份由监护人填写, 另一份由本人填写,青少年填写的问卷单独密封,然后放入更大的监护人调查问卷的信封里邮寄回复。
调研的主要方式是邮件调研问卷。收集信息内容包括:
① 各个年龄阶段针对性问题:如新生儿调查内容广泛,包括家庭、儿童保育、养育子女、健康、生活方式和社会经济地位;每年调查问卷都有针对性,涵盖各个年龄段相关内容;
在幼儿阶段,重点是儿童保育、育儿政策、态度和经验;在小学和初中阶段, 调查广泛涉及与学校有关的主题,包括学习、朋友和儿童放学后的安排;
② 人体测量学数据,如身高、体重;
③ 健康和患病住院情况,覆盖20多种慢性疾病和传染病、伤害和其它健康问题。
主要研究结果和发现:
① 在第一次分娩时离开全职工作的妇女和育儿假期后重返工作岗位的妇女很可能生第二个孩子。第一个孩子出生后,父亲频繁分担家务和抚养子女的家庭,生育第二个孩子的概率增加。此外,焦虑程度高、育儿负担重的女性想生第二个孩子的概率小。
② 母乳喂养对健康结果有积极的影响。纯母乳喂养6个月可降低早期哮喘发生率和呼吸道感染住院率。婴儿时期的纯母乳喂养和部分母乳喂养对儿童时期超重和肥胖有保护作用,男孩尤其如此;
纯母乳喂养和部分母乳喂养对幼儿从事适合年龄的行为的能力产生了积极影响;但同时也发现,与配方奶粉喂养相比,完全和部分母乳喂养婴儿到30个月龄时罹患蛀牙的风险更高。
③ 父母吸烟促进儿童哮喘发生。母亲在室内吸烟增加了婴幼儿哮喘风险,母父亲同时吸烟的家庭(无论室内或室外),其儿童因哮喘住院风险显著增加。然而,那些调查一开始父母就吸烟的儿童,其哮喘发生与父母亲戒烟无关。
(七)中国大陆出生队列研究
我国出生队列研究始于10年前,相比大部分北欧国家(1996-2001年)而言起步不算晚,发展迅速;但是我国大陆各地出生队列在设计、规模和实施内容上不尽相同。以下简介我国大陆安徽、上海、广州的出生队列研究。
1、中国-安徽出生队列(China-Anhui Birth Cohort Study, C-ABCS)
中国-安徽出生队列的目的是研究相关不良妊娠结局发生的原因和机制,全面促进优生优育工作。
该研究纳入安徽省6个地区2008年11月到2010年10月的16,776名孕妇参与调研随访(入组率93.8%),共收集13,454名单胎新生儿;研究分三阶段进行队列随访,第一阶段主要包括孕早期、中期和晚期的问卷调查以及血清样本收集,了解各类生物、物理、化学、营养、心理等环境暴露因素,以及生活方式、家庭环境、妊娠并发症等信息。
儿童出生后随访至1周岁,分别在42天、3月龄、6月龄、9月龄及12月龄进行健康体检、收集出生缺陷、生长发育登记资料。
第二阶段计划在儿童1.5-6岁收集饮食模式、养育方式、气质、行为、智力发育状况等信息;3-6岁学龄前儿童,发现多种孕期暴露与学龄前儿童超重/肥胖、行为问题、执行功能等方面的相关性;第三阶段主要调查8-15岁青春发动时相、体内性激素水平的变化状况。
中国-安徽出生队列重点研究生命早期环境暴露与出生缺陷及儿童发育异常的病因关联,该队列报道了迄今为止我国最为接近真实水平的、以社区为基础的总出生缺陷检出率(46.1%),32.7%为结构性出生缺陷,15.5%为功能性出生缺陷;
建立了孕妇血清双酚A、邻苯二甲酸酯、镉、锌、维生素D等物质的检测方法,发现多种与不良妊娠结局、出生缺陷及儿童发育异常有关的环境暴露因素。
2、广州出生队列(Born in Guangzhou Cohort Study, BIGCS)
广州出生队列研究将微生物与疾病之间的联系作为重点调查对象,收集孕期母亲接触的环境、生物、社会因素对胎儿生长发育的影响,并观察这些因素对儿童健康的长期影响,从而验证一系列不良妊娠结局和儿童期疾病的病因假设。
该研究于2010年开始筹建,2012年3月正式启动,招募对象是在广州市妇女儿童医学中心产检及分娩,并在广州长期居住的孕妇,随访从出生后到成年期。
截至2016年4月已招募17,480名孕妇入组,其中分娩13,481人、双胎302对、三胎1例,回收了75,422份问卷,105,300份生物血样本,计划到2020年招募3万人。
使用微博、QQ、短信平台等多种方式管理队列,建立围绕出生缺陷、妊娠期糖尿病和多肽儿童建立为主题的亚队列,孕期收集母亲血液、尿液和粪便,分娩时收集脐带血及胎盘、婴儿期收集纸血片、粪便、血液,儿童期收集血液、口腔颊粘膜脱落细胞及粪便。
目前主要研究成果包括孕期膳食、孕酮药物使用、气温变化等儿童健康的影响,以及广州市出生体重与全球标准的比较。
3、上海优生儿童队列(Shanghai Birth Cohort, SBC)
2012年启动的上海优生儿童前瞻性队列研究的目的是重点研究环境、遗传和行为因素对女性生殖健康、妊娠结局、儿童生长发育及疾病发生的影响,找到疾病的早期标志物,或“易感因素”,进而通过干预,改善疾病的发生发展。
该项研究由上海交通大学附属新华医院环境与儿童健康教育部、上海是重点实验室负责,在上海市5家医疗机构参与研究,该出生队列由孕产妇队列和儿童队列两部分组成,计划在上海妇幼保健机构招募3,000多名孕前及孕早期妇女作为调查对象。
孕前妇女通过两月一次的电话调查跟踪随访15个月,孕妇则分别在孕早期、中期及晚期在医院进行问卷调查,收集孕期营养、心理状态,采集血液、尿液等生物样本,挑选队列中30%的孕妇进行上门随访,开展家庭环境及营养状况评估、家庭环境采样的现场工作。
儿童出生时收集血清和胎盘样本,分别于出生后42天、6月龄、12月龄、24月龄随访儿童,进行喂养状况调查、生长发育评估、智力测试、自闭症筛查、行为习惯调查等工作。
新营养留言互动
您的电话不会被公开。 必填项已用 * 标注
相关热词搜索:
上一篇:一个方案给你新品成功的四大砝码,功能油脂浓缩乳液或将引爆新一轮市场
下一篇:橙汁会成为继咖啡之后中国市场最具潜力的品类吗?答案即刻揭晓
新营养峰会/活动


新营养热门资讯

新营养周报 | 农夫山泉上新蓝靛果混合汁
数据 / 观察 · 01-06

新营养周报 | 蒙牛推出早餐奶新品、纽曼
数据 / 观察 · 12-29

一图读懂 | 2024新营养肠道微生态格局与
XINGRAPHIC · 12-29

新营养周报 | 达芬奇优形推出纤体杯新品
数据 / 观察 · 12-23
新营养合作伙伴